Clear Sky Science · tr
Pro-β-karboksizom montajında biyomoleküler kondensat oluşum aşamaları
Neden küçük karbon fabrikaları önemli
Dünya’nın besin zincirleri fotosenteze dayanır, ancak havadan karbondioksiti (CO2) çeken ana enzim Rubisco yavaştır ve oksijen tarafından kolayca yanılabilir. Siyanobakteriler — mikroskobik, fotosentetik mikroorganizmalar — bu sorunu uzun zaman önce Rubisco etrafında CO2’yi hapseden protein “mikro-fabrikaları” olan karboksizomları inşa ederek çözdü. Bu çalışma, ApN ve CM adı verilen proteinlerden oluşan ve çalışır bir karboksizomun inşasına yardımcı olan kritik bir adaptör kompleksinin, kendiliğinden aşamalı olarak nasıl bir araya geldiğini çözümlüyor. Bu koreografiyi anlamak, benzer CO2-artırıcı sistemleri mahsul bitkilere yerleştirme ve verimi artırma çabalarına yol gösterebilir.
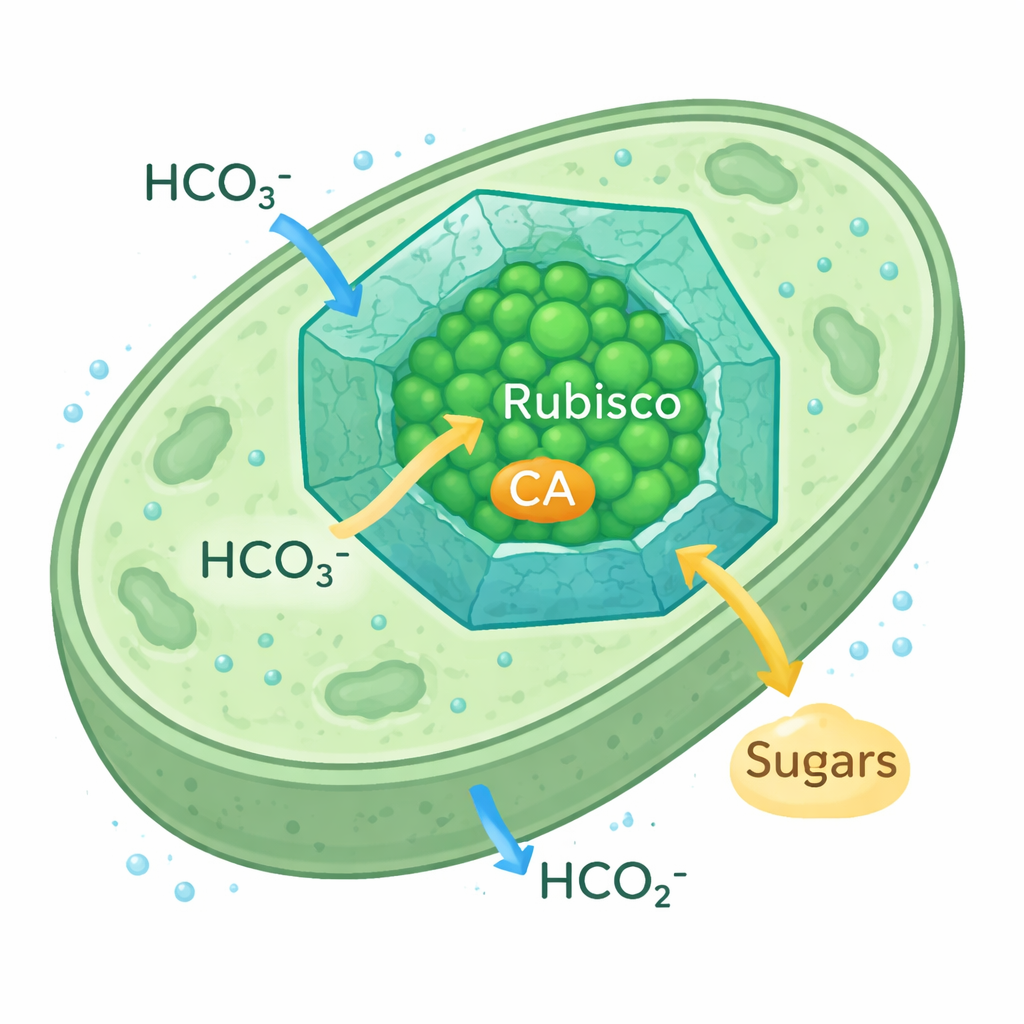
Microplar karbon yakalamayı nasıl güçlendirir
Siyanobakteriyel karboksizomlar hücre içinde yer alan, küçük, ikozahedral protein bölmeleri gibidir. Protein kabuk, bikarbonat iyonlarının girişine izin verirken CO2’nin kaçışını yavaşlatarak Rubisco ve bikarbonatı CO2’ye dönüştüren yardımcı enzim karboanhidrazı (CA) çevreleyen CO2 açısından zengin bir baloncuk oluşturur. Bu CO2 yoğunlaştırma mekanizması, siyanobakterilerin çevredeki CO2 az olsa bile başarılı olmasını sağlar. Bu karboksizomlarda kullanılan Rubisco biçimi bitki Rubisco’suna benzer olduğu için araştırmacılar, bitki kloroplastlarını daha verimli karbon yakalayacak şekilde yeniden mühendislik etmek için gerçekçi bir model olarak görüyorlar.
Kabuğa bağlanan puzzle parçası
Katı protein kabuk oluşmadan önce, siyanobakteriler önce yumuşak, damla benzeri bir “pro-karboksizom” kondensatı inşa eder. Önceki çalışmalar, CM adlı bir iskelet proteininin Rubisco ve CA’yı bu damla içinde bir araya getirdiğini göstermişti. Eksik halkaysa kabuk adaptör proteini ApN’in bu yapıya nasıl katıldığı ve daha sonra kabuk proteinlerine nasıl bağlandığıydı. Yazarlar, ApN tek başına üretildiğinde diğer karboksizom bileşenlerini görmezden gelen dört parçalı (tetramer) bir demet oluşturduğunu buldular. Bu durumda ApN, Rubisco, CM ve CA ile damlalara çekilemiyordu; bu da gerçek karboksizom montajı sırasında farklı bir ApN formunun kullanılması gerektiğini gösteriyor.
Birlikte inşa edilen adaptörler ve bir redoks anahtarı
Siyanobakterilerdeki genetik düzenlemeye bakıldığında ekip, apn geninin aynı operonda cm’nin hemen arkasında yer aldığını ve bu iki proteinin genellikle birlikte üretildiğini fark etti. Bunu bakterilerde taklit ederek, ortak bir DNA parçasından ApN ve CM’yi birlikte ifade ettiler. Bu koşullarda ApN artık kendi başına bir tetramer oluşturmadı. Bunun yerine üç ApN alt birimi ve bir CM alt birimi, (ApN)3:CM olarak gösterilen spesifik dört parçalı bir adaptör kompleksi halinde bir araya geldi. Bu hetero-kompleks Rubisco, CA ve CM’ye kolayca bağlandı ve kondensatlara katılabildi. Yapısal modelleme ve kriyo-elektron mikroskobu, ApN ile CM’nin ince ayarlı bir “dil ve oluk” arayüzü aracılığıyla buluştuğunu gösterdi; arayüzdeki kilit ApN kalıntıları mutasyona uğratıldığında adaptör oluşmadı ve hücreler düzgün karboksizomlar oluşturamadı.
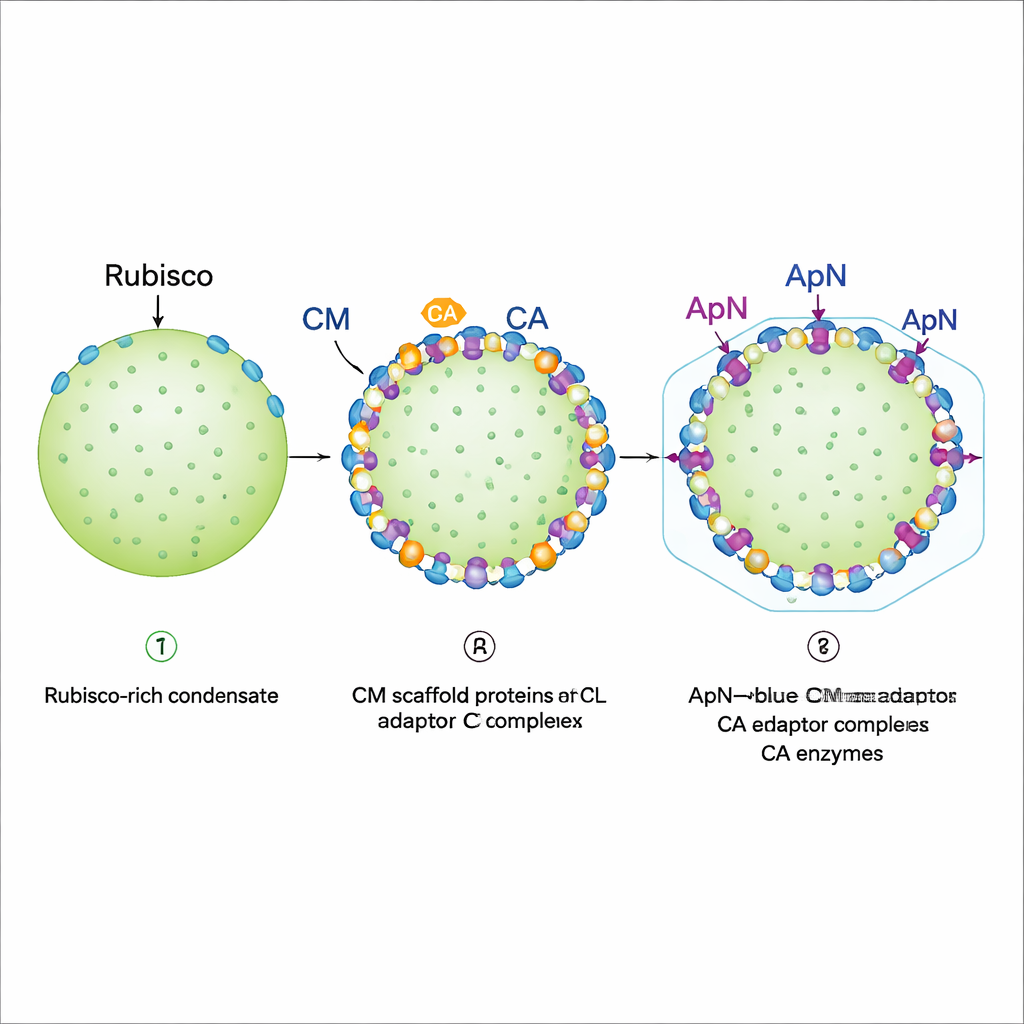
Kenar konumlandırması ve kabuk için hazırlık
Yeniden oluşturulmuş damlaların floresan görüntülenmesi adım adım bir montaj programını ortaya koydu. Önce Rubisco, tam boy CM ve daha kısa bir CM varyantı yoğun, jel benzeri davranan bir kondensat oluşturuyor: Rubisco pratikte yerinde kilitlenmiş durumda. Ardından CA ve (ApN)3:CM kompleksi ekleniyor. Her ikisi de, CA (ApN)3:CM’den önce veya onunla birlikte geldiği sürece mevcut damlanın dış kenarında yerleşmeyi tercih ediyor. Bu kenar yerleşimi önemlidir çünkü ApN’in kuyruğu kabuk proteinlerine bağlanan bir “kapsülleme peptidi” taşır. Yazarlar ayrıca yerleşik bir redoks anahtarı keşfettiler: olgunlaşan karboksizomun iç kısmı daha oksitleyici hale geldikçe, ApN’deki iki korunan sistein kalıntısı oksitlenebiliyor ve adaptörü (ApN)3:CM tetramerinden (ApN)2:CM trimere doğru itilim yapabiliyor. Canlı siyanobakteriyel hücrelerde bu sisteinleri değiştirmek büyümeyi yavaşlattı ve hâlâ işlevsel olmakla birlikte aşırı büyük karboksizomların oluşmasına neden oldu; bu da olgunlaşmayı incelikle ayarlamadaki rollerini vurguluyor.
Daha iyi bitkiler inşa etmek için dersler
Uzman olmayanlar için bu moleküler ayrıntılar arkaik gelebilir, ancak öz basittir: zamanlama ve ortaklık önemlidir. Çalışma, ApN’in işe yaramaz kendi kendine kümeler oluşturmaktan kaçınmak ve onun yerine pro-karboksizom yüzeyine park eden, kabuğu işe almaya hazır bir adaptör haline gelmek için CM ile birlikte üretilmesi gerektiğini gösteriyor. Ardından bir redoks-duyarlı adım, bölme kapanırken ve iç kimyası değişirken bu adaptörü yeniden şekillendirmeye yardımcı oluyor. Cyanobakteriyel CO2 yoğunlaştırma mekanizmasını mahsullere aktarmayı uman bitki mühendisleri için çalışma, tek tek parçaları taşımaktan daha fazlasının gerektiğini; doğal karboksizom montajını yönlendiren sıra, oranlar ve kimyasal tetikleyicilere de saygı gösterilmesi gerektiğini vurguluyor.
Atıf: Zang, K., Hong, X., Nguyen, N.D. et al. Stages of biomolecular condensate formation in pro-β-carboxysome assembly. Nat. Plants 12, 447–464 (2026). https://doi.org/10.1038/s41477-026-02227-6
Anahtar kelimeler: karboksizom montajı, siyanobakteriyel CO2 yoğunlaştırma mekanizması, Rubisco kondansatları, biyomoleküler kondensatlar, sentetik fotosentez