Clear Sky Science · tr
Mycobacterium tuberculosis’ta transkripsiyon başlatımında duraklamanın yapısal temeli
Bir mikroorganizmanın gen anahtarları nasıl takılabilir
Verem etkeni Mycobacterium tuberculosis, vücudun zorlu ortamında hayatta kalmak için genleri sürekli olarak açıp kapatmak zorundadır. Bu makalede anlatılan çalışma, bu genetik kontrolde en erken adımlardan birine bakıyor ve hücrenin DNA’yı RNA’ya çevirmeye başladığı anda oluşan kısa süreli bir “duraklamayı” yakalıyor. Bu duraklamayı anlamak, mikroorganizmanın bir geni tamamen açıp açmamaya veya denemeyi sonlandırıp kısa RNA parçalarını serbest bırakmaya nasıl karar verdiğini açıklamaya yardımcı oluyor ve nihayetinde antibiyotikler için yeni zayıf noktaları ortaya çıkarabilir.
Mesajı DNA’dan Başlatmak
Bakterilerde RNA üretmenin ilk adımı olarak DNA boyunca ilerleyen büyük bir protein makinesi olan RNA polimeraz görev yapar. Başlamak için, onu DNA üzerindeki belirli başlangıç noktalarına yönlendiren ve çift sarmalı açarak küçük bir “kabarcık” oluşturmaya yardımcı olan sigma faktörleri olarak bilinen yardımcı proteinlerle ortak çalışır. Mycobacterium tuberculosis için sigma E adı verilen bu yardımcı faktör, bakterinin hücre zarının dışındaki stres altındayken özellikle önemlidir. Bir başka faktör olan CarD, açık DNA kabarcığını stabilize etmeye yardımcı olur. Birlikte, bu bileşenler başlatım kompleksi oluşturur; bu kompleks RNA’nın ilk birkaç yapı taşını yapar ve ardından ya tam uzunlukta bir mesaj için taahhüt eder ya da vazgeçip kısa RNA parçalarını bırakır.
Altı ya da Yedi Harf Uzunluğunda Gizli Bir Duraklama
Önceki biyokimyasal ve tek molekül çalışmalar, RNA ipliği yalnızca altı veya yedi yapı taşı uzunluğuna ulaştığında RNA polimerazın çok erken aşamada sık sık durakladığına dair ipuçları vermişti. Ancak bu duraklamalar son derece kısa sürer ve doğrudan gözlemlenmesi zordur; bu yüzden yüksek çözünürlüklü yapılar mevcut değildi. Bu çalışmada yazarlar, M. tuberculosis’in başlatım mekanizmasının birden çok çeşidini dondurup görselleştirmek için kriyo-elektron mikroskopisini kullandılar. RNA uzunluğu altı veya yedi birim olduğunda kompleksi yakalayan DNA–RNA iskeleleri tasarladılar ve üç ana durumun yapısını çözdüler: iki “düzenli” başlatım kompleksi ve ayrı bir duraklamış başlatım kompleksi.
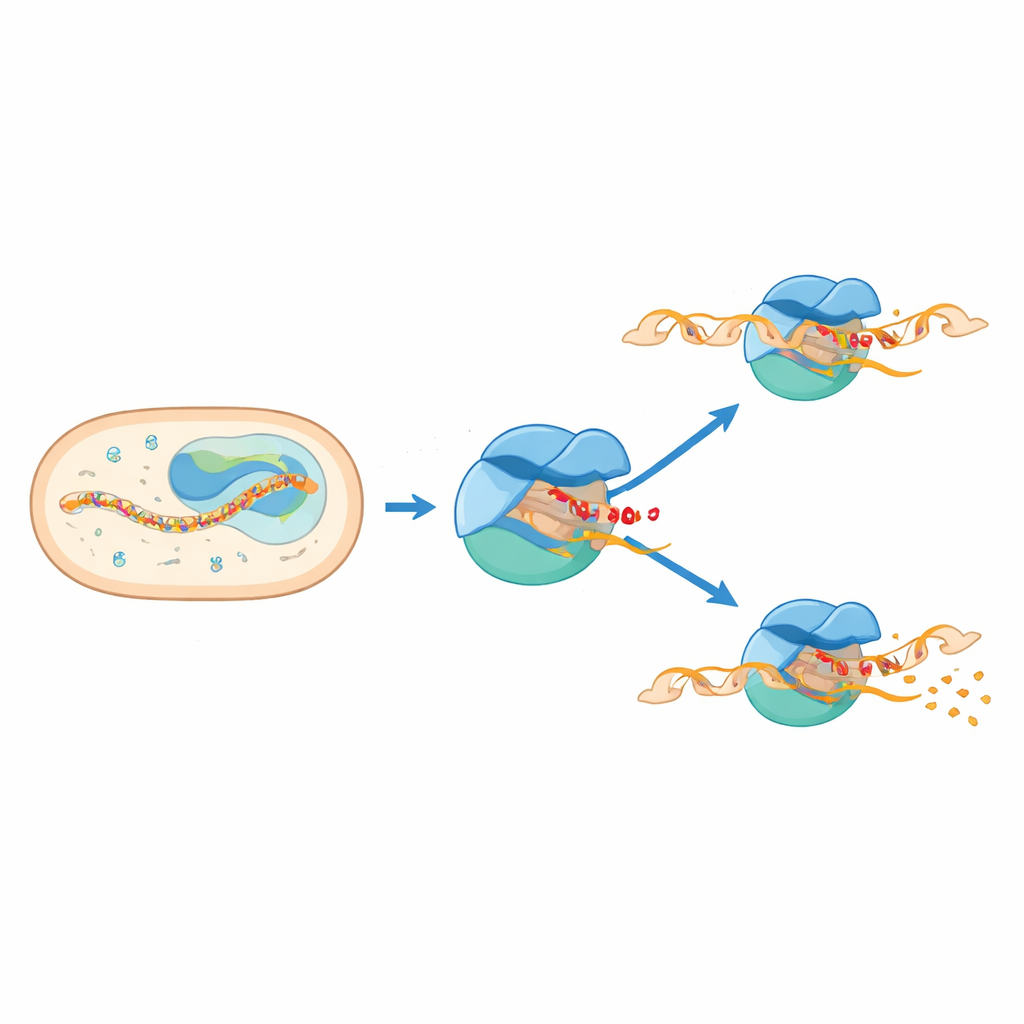
Makinanın İki Parçası Çarpıştığında
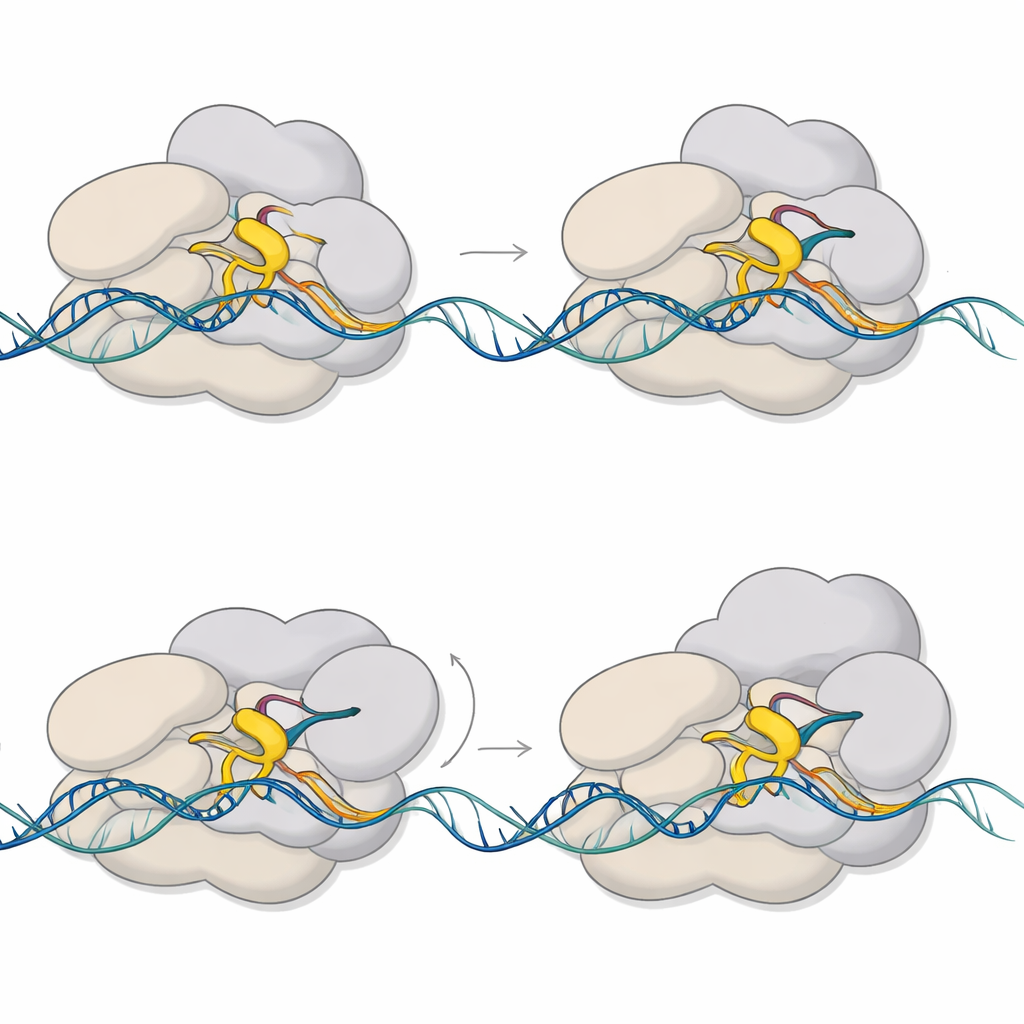
Yapılar, duraklamanın mekanizmasının makinenin iki parçasının aynı alanı işgal etmeye çalışmasından kaynaklandığını ortaya koyuyor. Sigma E’nin esnek bir bölgesi olan 3.2 bölgesi, büyüyen RNA’nın enzimden çıkarken geçmesi gereken kanala doğru uzanır. RNA altı ya da yedi birime uzadıkça ön ucu bu çıkıntılı halka ile çarpışır. Bu çarpışma, RNA polimeraz içindeki büyük bir modülün dönmesine ve köprü heliksi olarak bilinen önemli bir yapısal elemanın deformasyona uğramasına neden olur. Aynı zamanda aktif bölgenin hemen yukarısındaki DNA içe doğru çekilerek kısmen çözülür; bu, “sıkışmış” ve bükülmüş bir kabarcık yaratır. Bu birbirine bağlı hareketler, kompleksi mekanik gerilim depolayan yarı-ilerlemiş, duraklamış bir duruma kilitler.
Şekil Değiştiren Bir Yardımcı Kabarcığı Sabitliyor
Aksesuar faktör CarD, önce düşünülenden daha çok yönlü bir role sahip çıkıyor. Duraklamamış başlatım kompleksinde CarD, kabarcığın kenarındaki bir DNA ipliğinin minör oluğuna girer ve RNA polimerazın senteze başlaması için kabarcığın açık kalmasına yardımcı olur. Duraklamış komplekste ise CarD konum değiştirerek şablon olmayan DNA ipliğini kavrar ve sigma E ile birlikte sıkışmış kabarcığı daha sıkı bir şekilde sıkıştırır. Biyokimyasal testler, CarD’nin sigma E’ye bağımlı transkripsiyonu artırdığını ve altı-yedi birim aşamasında biriken RNA molekül sayısını etkilediğini gösterir. Bu bulgular, CarD’nin hem ilk açık durumu stabilize ettiğini hem de duraklamış kompleksin erken çökmesini önlemeye yardımcı olduğunu düşündürür.
Başarısızlık ile Taahhüt Arasında Bir Kontrol Noktası
Kısmen önceden açılmış DNA üzerinde oluşturulmuş yüksek çözünürlüklü yapılarla, tamamen eşleşmiş DNA üzerinde bir araya getirilmiş daha düşük çözünürlüklü yapıları karşılaştırarak yazarlar, aynı temel duraklamış biçimlerin daha doğal koşullar altında da ortaya çıktığını gösteriyorlar. RNA’nın büyümesi ile sigma 3.2 halkası arasındaki çarpışmanın altı ila yedi RNA biriminde enerji yüklü bir kontrol noktası oluşturduğunu öne sürüyorlar. Bu hazır durumdan kompleks iki ana yoldan birini izleyebilir: kısa RNA’yı serbest bırakarak sıfırlanma—abortif bir deneme—veya depolanan gerilimi kullanarak promotör bölgesinden kurtulup istikrarlı, uzun süreli RNA üretimine girmek. CarD’nin yeniden konumlanması ve sigma faktörünün yukarı akıştaki DNA üzerindeki kavrayışının gevşemesi bu kararı yönlendiren etkenler gibi görünmektedir.
Bu Neden Verem Açısından Önemli
Genel okuyucu için çıkarılacak ana mesaj, verem bakterisinin genleri bir ışık anahtarı gibi basitçe açıp kapamadığıdır. Bunun yerine, hücre küçük bir RNA uzunluğunda tereddüt eder ve bu kısa mekanik duraklamayı abort etme ile tamamen açılma arasında bir karar noktası olarak kullanır. Bu çalışma o duraklamayı atomik ayrıntıda yakalıyor ve küçük hareketli bir halkacığın, dönen bir çekirdeğin ve yeniden şekillenmiş bir DNA kabarcığının birlikte nasıl kontrol edilebilir bir kontrol noktası yarattığını gösteriyor. Sigma E ve CarD M. tuberculosis’in strese yanıtında merkezi olduğu için bu bulgular, araştırıcıların bu erken adımı kilitleyen ve bakterinin uyum sağlama ve hayatta kalma yeteneğini köreltmeyi amaçlayan ilaçlar tasarlamasına nihayetinde yardımcı olabilir.
Atıf: Zheng, L., Xu, K. Structural basis of pausing during transcription initiation in mycobacterium tuberculosis. Nat Commun 17, 2197 (2026). https://doi.org/10.1038/s41467-026-69104-w
Anahtar kelimeler: transkripsiyon başlatımı, RNA polimeraz duraklaması, Mycobacterium tuberculosis, sigma faktörleri, cryo-EM