Clear Sky Science · tr
F-box helikaz 1 etrafında kurulu SCF ubiquitin ligaz kompleksinin çatal tersine çevirme ve RAD51 düzenlemesinin yapısal temeli
Hücreler tıkanan bir DNA kopyalama makinesini nasıl kurtarıyor
Her hücre bölünmesinde milyarlarca DNA harfini hızlı ve doğru şekilde kopyalamak zorundadır. Ancak bu kopyalama makinası sık sık sorunla karşılaşır—DNA hasarı, kopyalanması zor diziler veya ilaç tedavileri süreci durdurabilir. Böyle olduğunda hücreler, kopyalama çatallarını stabilize etmek ve yeniden başlatmak için acil onarım ekiplerine güvenir. Bu makale, bu ekiplerden birinin liderinin—helikaz FBH1 etrafında kurulan bir protein kompleksinin—tıkalı DNA’yı nasıl yeniden şekillendirdiğini ve başka bir önemli onarım proteini olan RAD51’i nasıl sıkı kontrol altında tuttuğunu eşi görülmemiş yapısal ayrıntıda ortaya koyuyor. Bu koreografiyi anlamak, hücrelerin tehlikeli genom kırılmalarından nasıl kaçındığını ve bu faktörlerdeki bozulmaların neden kansere bağlandığını açıklamaya yardımcı olur.
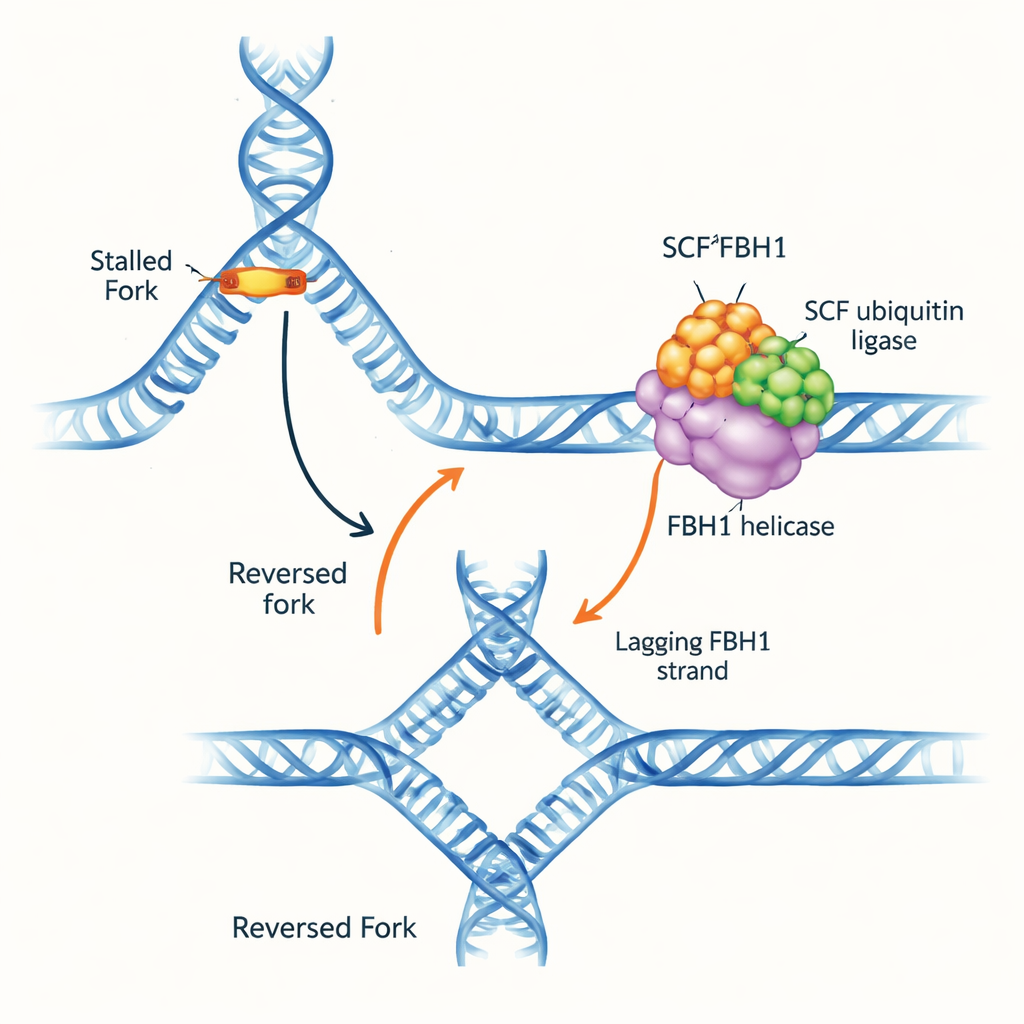
DNA otoyolunda bir trafik sıkışıklığı
DNA replikasyon çatalları, çift sarmalın açıldığı ve kopyalandığı Y biçimli yapılardır. Çatal engellerle karşılaştığında yönünü tersine çevirebilir: ebeveyn zincirler yeniden eşlenir ve yeni sentezlenen zincirler birbirleriyle eşleşir, dört kollu bir DNA kavşağı oluşur. Bu “çatal tersine çevirme” hasarı onarmak ve replikasyonu güvenli şekilde yeniden başlatmak için zaman kazandırabilir, ancak kötü yönetildiğinde kırılmaya yatkın DNA oluşturur. Birkaç moleküler motor, çatalları geri itmek için çatalın önündeki çift sarmal DNA’yı kavrayarak görev yapar. FBH1 ise farklı davrandığı için öne çıkıyordu ve tümörlerde sıkça değişikliğe uğraması, onun ayrı ve tıbbi açıdan önemli bir yolla çalıştığını düşündürüyor.
Belirli bir yanağı tercih eden çatal uzmanı
Yazarlar, FBH1 helikazını ubiquitin etiketleme modülüyle (SCF) birleştiren insan SCFFBH1 kompleksini saflaştırdılar. Önce FBH1’in hangi DNA biçimlerini tercih ettiğini sordular. Bağlanma ve açma deneyleriyle, SCFFBH1’in gerçek çatal benzeri yapılara, özellikle “geciken” kalıp zincirinde—küçük parçalar halinde kopyalanan taraf—kısa bir tek iplikçikli DNA bölgesi bulunduğunda kuvvetle tercih gösterdiğini gösterdiler. Bu yapılarda FBH1, basit bir serbest uç üzerinde olduğundan yüzlerce kat daha hızlı DNA açıyor; bu da çatal kavşağında doğru yerleşimin motor aktivitesini aşırı hızlandırdığını gösteriyor. Tek molekül manyetik forseps deneyleri, kompleksin tek iplikli DNA boyunca düşmeden binlerce baz taşıyabilecek şekilde açma yapıp daha sonra ilerleyebildiğini doğruladı; yani güçlü, prosessif bir motor gibi davranıyor.
Arkadan çekiş ile çatalı tersine çevirme
Bu motorun çatalı aslında nasıl tersine çevirdiğini bulmak için ekip, tek iplikçikli boşluklu veya boşluksuz yapay çatal alt tabakalarını test etti. Diğer çatal yeniden şekillendiricilerden farklı olarak SCFFBH1 tam eşlenmiş bir çatal üzerinde hareket edemedi; geciken kalıp üzerinde bir boşluk gerekiyordu ve 3′–5′ yönünde o tek iplik boyundan ilerliyordu. Bu “arkadan çekiş” hareketi, enzimin çatal kavşağında sıkıca tutunurken ebeveyn zincirlerin yeniden eşleşmesini sağlıyor. Tıkalı bir çatala bağlı SCFFBH1’in bir kriyo-elektron mikroskobu yapısı bunun nasıl olduğunu gösteriyor: FBH1’in bir alt bölgesindeki özel pozitif yüklü bir yamaya, bir “kavşak bağlama motifi”ne, üç DNA kolunun buluştuğu dal noktası tamında yuva gibi oturuyor. Araştırmacılar bu motifteki iki ana amino asidi mutasyona uğrattıklarında, test tüplerinde çatal tersine çevirme yaklaşık 25 kat azaldı; buna karşın temel açma motoru hâlâ makul düzeyde çalışıyordu. Hücrelerde aynı mutasyon, stres oluşturan bir ilaca yanıt olarak replikasyon çatallarının normal olarak yavaşlamasını engelledi; bu da başarısız çatal tersine çevirmenin ayırt edici bir işaretiydi.
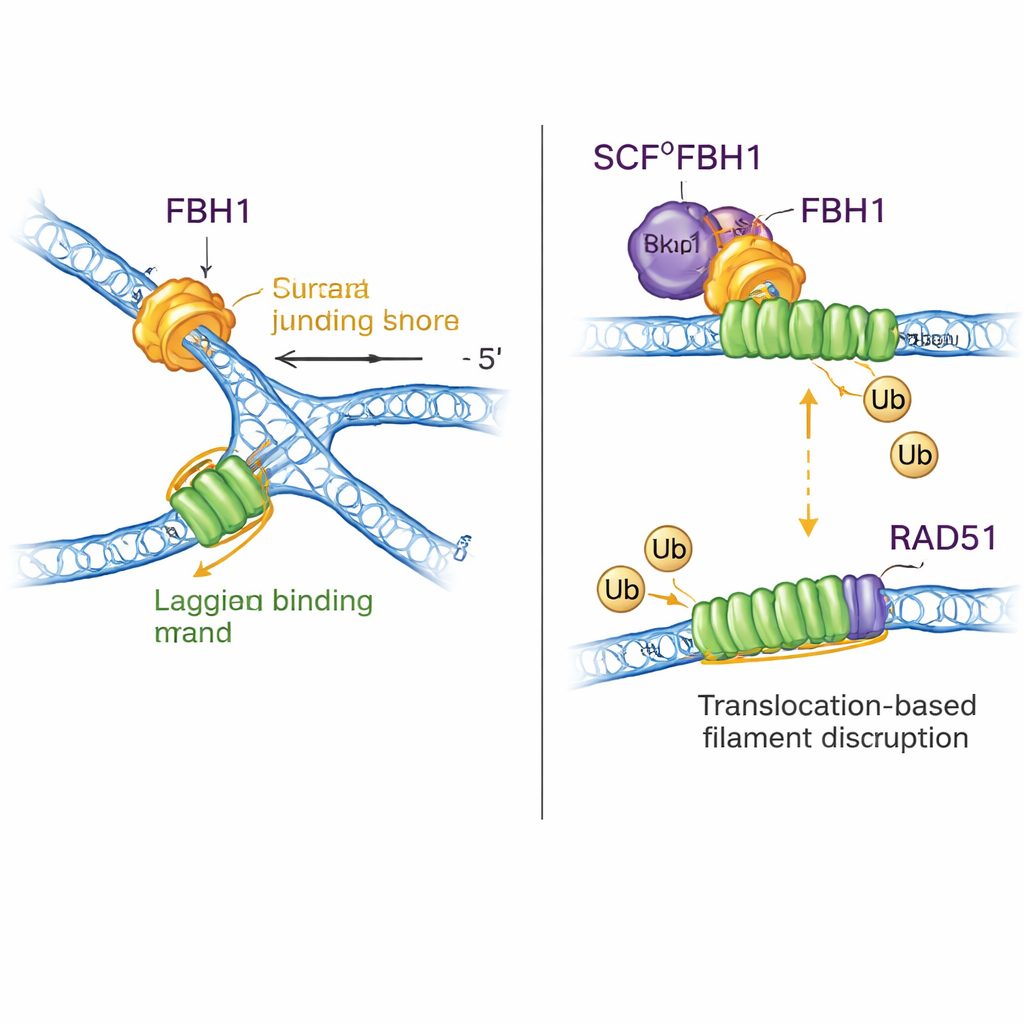
Çifte rol: DNA’yı yeniden şekillendirme ve RAD51’i uzaklaştırma
FBH1 sadece bir motor değil; SCF kompleksi aracılığıyla proteinlere küçük ubiquitin etiketleri takılmasına da yardımcı olur. Yeni yapılar, SCFFBH1–DNA kompleksinin neredeyse tamamını bir araya getiriyor ve ligazın ubiquitin transfer “kafasının”, FBH1’den çıkan geciken zincir DNA’sından 45 Angstromdan az bir uzaklıkta oturduğunu gösteriyor. Diğer ubiquitin ligazlarının bilinen yapılarının kenetlenmesiyle yazarlar, bu geometrinin RAD51’i—DNA üzerinde filamentler oluşturarak rekombinasyonu teşvik eden bir enzimi—tıkalı bir çatalın geciken ipliği üzerinde hedef hattına yerleştirdiğini öne sürüyor. Önceki çalışmalar FBH1’in RAD51’i DNA’dan mekanik olarak sökebildiğini ve ayrıca onun kolayca yeniden bağlanmaması için ubiquitinle işaretleyebildiğini göstermişti. Yeni model bu fikirleri birleştiriyor: FBH1 geciken ipliği boyunca çekiş yaparken çatalı yeniden şekillendirirken, yakınlardaki SCF modülü RAD51 alt birimlerini etiketleyerek aşırı RAD51 filamentlerinin parçalanmasına veya yeniden oluşmasının önlenmesine yardımcı oluyor; bu filamentler aksi takdirde uygun çatal işlenmesini engelleyebilir.
Genom kararlılığı ve kanser için anlamı
Bir arada ele alındığında, çalışma SCFFBH1’in tıkalı DNA replikasyon çatallarını diğer bilinen çatal yeniden şekillendiricilerden farklı bir mekanizmayla tersine çevirdiğini gösteriyor: çatal kavşağını sıkıca kavrarken arkadan geciken kalıp üzerinde yer değiştirerek ebeveyn DNA’nın güvenli şekilde yeniden sarmalanmasını teşvik ediyor; uzun ömürlü hareketli bir dört kollu kavşak oluşturmuyor. Aynı zamanda, içine yerleşmiş ubiquitin ligazı, aynı iplikten RAD51’i uzaklaştırmak için stratejik olarak konumlanmış durumda; bu, replikasyon stresi sırasında kontrolsüz rekombinasyonu önlüyor. FBH1’in kanserlerde sıkça kaybolması veya mutasyona uğraması nedeniyle, bu yapısal ve mekanistik bulgular bu tür tümörlerin neden DNA hasarı biriktirdiğini ve replikasyon stresini tetikleyen ya da ilişkili yolları hedef alan ilaçlara neden farklı yanıt verebileceklerini açıklamaya yardımcı oluyor.
Atıf: Greer, B.H., Mendia-Garcia, J., Mullins, E.A. et al. Structural basis for fork reversal and RAD51 regulation by the SCF ubiquitin ligase complex of F-box helicase 1. Nat Commun 17, 2041 (2026). https://doi.org/10.1038/s41467-026-68752-2
Anahtar kelimeler: DNA replikasyon stresi, çatal tersine çevirme, FBH1 helikazı, RAD51 düzenlemesi, genom kararlılığı