Clear Sky Science · tr
Caenorhabditis elegans’te DNA metilasyonu olmayan bir organizmada MBD‑2'nin NuRD ile birlikte ve ayrıksı olarak çift genomik yerleşimi ve gen düzenleyici işlevleri
Küçük solucanlar gen kontrol kurallarını nasıl yeniden yazar
Hücrelerimiz, hangi genlerin açık ya da kapalı tutulacağını hatırlamak için DNA ve proteinler üzerinde kimyasal işaretler kullanır. En bilinen işaretlerden biri olan DNA metilasyonu, bazı hayvanlarda yoktur—yine de bu hayvanlar normal şekilde gelişir ve ürer. Bu makale, küçük bir solucan olan Caenorhabditis elegans'ın DNA metilasyonu olmadan gen kontrolünü nasıl sağladığını inceliyor; beklenmedik derecede esnek bir yedek sistem ortaya koyuyor ve epigenetik konusundaki düşüncelerimizi yeniden şekillendirebilir.
Eksik bir DNA işareti büyük bir soru doğuruyor
Memelilerde 5‑metilsitozin (5mC) adlı bir kimyasal işaret, gelişim sırasında genlerin susturulmasına, X kromozomunun inaktivasyonuna ve hareketli genlere karşı savunmaya yardımcı olur. MBD2 ve MBD3 adı verilen proteinler bu işaretleri okur ve NuRD kompleksi adı verilen büyük bir protein makinesini çekerek kromatinin yeniden şekillenmesine ve genlerin kapatılmasına aracılık eder. Garip bir şekilde, C. elegans ve meyve sinekleri de dahil olmak üzere birçok omurgasız, 5mC'yi ve onu yazan enzimleri kaybetmiş olsa da MBD2/3 benzeri bir proteini korumuştur. Bu durum bir bilmece ortaya çıkarıyor: DNA metilasyonunu artık kullanmayan bir genomda neden bir “DNA metilasyonu okuyucusu” tutulmuş ve bunun yerine ne yapıyor?
Solucanların onsuz yaşayamadığı bir iskele proteini
Yazarlar bu proteinin solucan versiyonu MBD‑2 üzerine odaklandı. Memelilerdeki akrabalarının aksine, solucan MBD‑2 klasik 5mC'yi tanıyan metil‑bağlayıcı domaini kaybetmiş, ancak diğer proteinlere tutunmaya uygun esnek ve sarmal bölgeleri korumuştur. MBD‑2'yi floresan işaretlerle etiketleyerek ekip, proteinin solucanın yaşamı boyunca hemen hemen tüm hücrelerin çekirdeklerinde bulunduğunu ve geniş bir gen düzenleme rolüne işaret ettiğini gösterdi. Genetik mühendislikle yazarlar, tamamıyla MBD‑2'siz veya yalnızca NuRD ile temas kuran sarmal‑kıvrım (coiled‑coil) bölgesi eksik solucanlar yarattı. Her iki mutasyon da ciddi sorunlara yol açtı: hayvanlar küçük, hareketleri zayıf, üreme yapıları bozuk gelişmiş ve büyük ölçüde kısırdı. Bu sonuçlar, DNA metilasyonu olmasa bile MBD‑2'nin normal gelişim ve üreme için zorunlu olduğunu ortaya koydu.
Metilasyonsuz dünyada NuRD makinesini yeniden kurmak
Solucan MBD‑2'nin hâlâ NuRD ile birlikte çalışıp çalışmadığını görmek için yazarlar, etiketlenmiş MBD‑2'yi solucan özütlerinden çekip ortaklarını kütle spektrometrisiyle tanımladı. Bilinen NuRD bileşenlerinin çoğu bulundu ve bu, MBD‑2'nin memelilerde MBD2/3'ün yaptığı gibi bu kompleksin temel bir iskelesi olarak işlev gördüğünü doğruladı. Sarmal‑kıvrım bölgesi silindiğinde, özellikle nükleozomları yeniden şekillendiren proteinlerle birçok etkileşim kayboldu. Aynı zamanda, geniş ölçekli RNA dizileme, MBD‑2 mutantlarında solucak genlerinin dörtte birinden fazlasının aktivitesinin değiştiğini, açılan gen sayısının kapananlardan çok daha fazla olduğunu gösterdi. Bu desen, MBD‑2'nin sıklıkla NuRD ile birlikte uygunsuz genleri susturmaya yönelik bir baskılayıcı olarak davrandığını, aynı zamanda yüksek düzeyde ifade edilen bir gen alt kümesinin uygun aktivitesinin sürdürülmesine yardımcı olduğunu işaret ediyor.
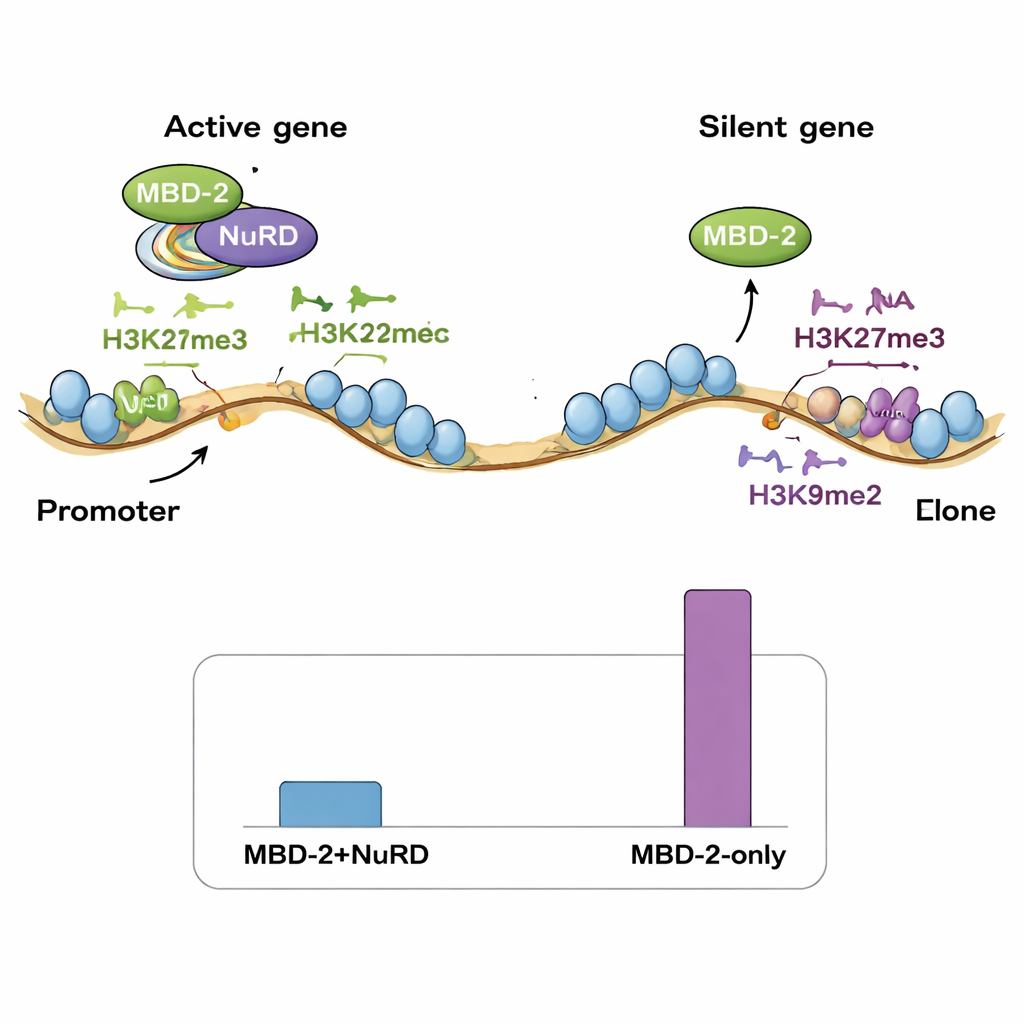
Genom üzerinde iki bağlanma modu
Ardından ekip, MBD‑2'nin genom boyunca nerelere oturduğunu ChIP‑seq ile haritaladı ve bu yerleri NuRD bileşenleri ve çeşitli histon işaretlerinin mevcut haritalarıyla karşılaştırdı. Şaşırtıcı biçimde, MBD‑2 bölgelerinin yalnızca küçük bir kısmı NuRD proteinleri ve açık, aktif kromatin işaretleri ile sıkı örtüşme gösterdi. MBD‑2 bölgelerinin büyük çoğunluğu ise H3K27me3 ve H3K9me2/3 gibi baskılayıcı histon işaretleri bakımından zengin kromozomal “kollar”da yer alıyordu. Başka bir deyişle, MBD‑2 çift davranış sergiliyor: azınlıktaki bazı bölgelerde NuRD ile birlikte aktif genleri ince ayar yapmak için hareket ederken, çok daha fazla bölgede zaten sessiz olan alanlara bağımsız olarak bağlanıyor. Gen içindeki bağlanma deseni de önem taşıyor—MBD‑2 gen başlangıç bölgelerine yakın kümelendiğinde proteinin kaybı bu genleri aktive etme eğilimindeyken, gen gövdeleri içinde daha derin bağlanma MBD‑2 kaldırıldığında sıkça ekspresyon azalmasıyla ilişkilendiriliyor.
Eksik DNA metilasyonu için evrimsel bir yedek
Yazarlar, DNA metilasyonunu kaybeden türlerde baskılayıcı histon işaretlerinin—özellikle H3K27me3'ün—benzer bir düzenleyici nişi doldurmak için genişlemiş olabileceğini öneriyor. Solucan MBD‑2, 5mC'yi DNA üzerinde okumak yerine şimdi belirli histon modifikasyonlarıyla işaretlenmiş kromatin bölgelerine bağlanmaya yönlendirilmiş gibi görünüyor; aynı zamanda korunmuş protein‑etkileşim domainleri aracılığıyla NuRD kompleksini hâlâ sabitliyor. Bu çalışma, eski bir gen‑susturma aracının evrim boyunca yeniden kablolanabileceğini gösteriyor: DNA üzerindeki kimyasal işaret kaybolabilir, ancak ilişkili protein makineleri alternatif sinyallere geçiş yaparak hayatta kalabilir. Uzman olmayanlar için ana mesaj şudur: gen düzenlemesi son derece uyarlanabilir—hücreler büyük bir epigenetik işareti kaybedebilir ve buna rağmen diğer kromatin etiketlerine ve MBD‑2 gibi çok yönlü iskelesel proteinlerin rollerine daha fazla dayanarak binlerce genin karmaşık kontrolünü koruyabilir.
Atıf: Tsui, H.N., Wong, C.Y.Y., Zheng, C. et al. Dual genomic localizations and gene regulatory functions of MBD-2 with and without NuRD in Caenorhabditis elegans which lacks DNA methylation. Nat Commun 17, 1875 (2026). https://doi.org/10.1038/s41467-026-68592-0
Anahtar kelimeler: epigenetik, kromatin, gen düzenlemesi, C. elegans, histon modifikasyonu