Clear Sky Science · sv
Steg i bildandet av biomolekylära kondensat vid pro-β-karboxysom‑sammansättning
Varför små kolfabriker spelar roll
Jordens näringskedjor är beroende av fotosyntes, men det centrala enzymet som fångar koldioxid (CO2) från luften, Rubisco, är långsamt och lätt förväxlas av syre. Cyanobakterier – mikroskopiska, fotosyntetiska mikrober – löste detta tidigt genom att bygga proteinbaserade ”mikrofabriker” kallade karboxysomer som fångar CO2 kring Rubisco. Denna studie nystar upp hur ett avgörande adapterkomplex, bestående av proteinerna ApN och CM, själv‑sammansätts i steg för att hjälpa till att bygga en fungerande karboxysom. Att förstå denna koreografi kan vägleda försök att installera liknande CO2‑förstärkande system i grödor och därigenom öka avkastningen.
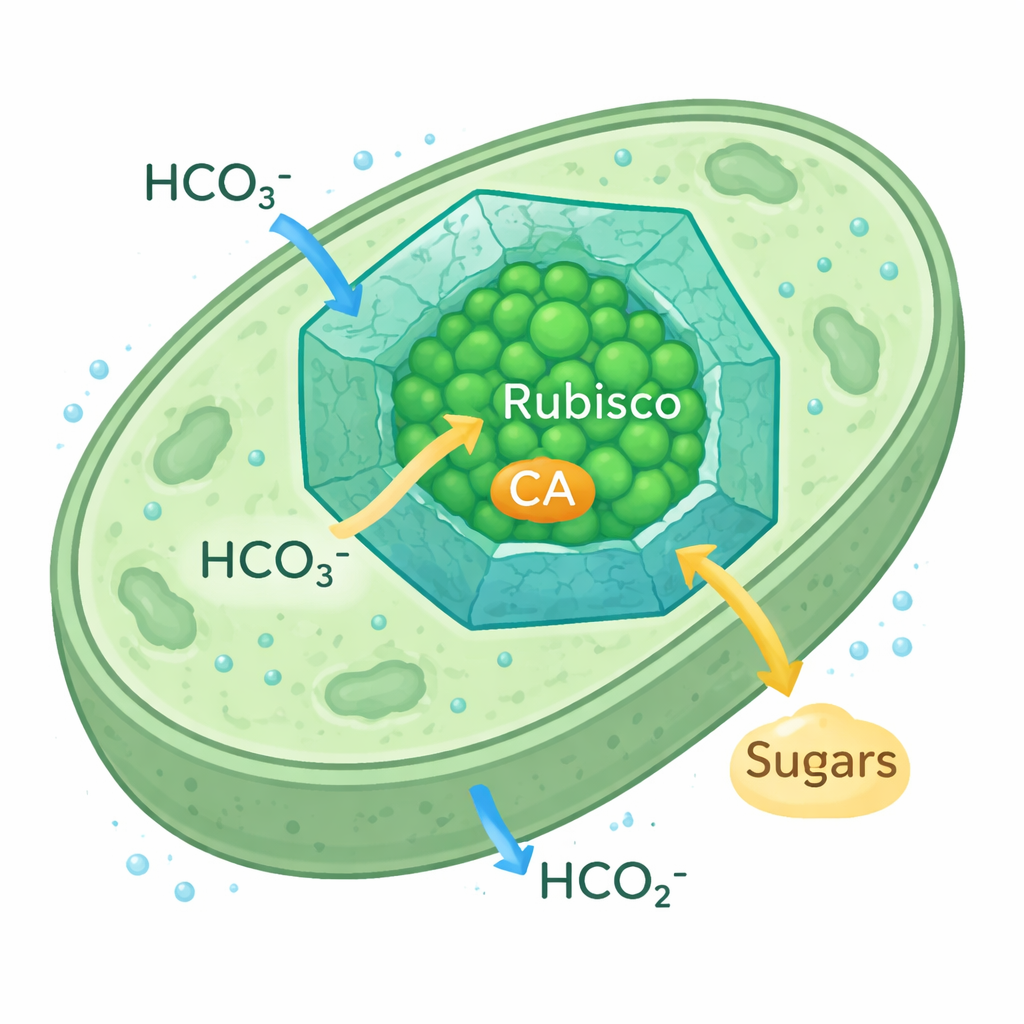
Hur mikrober överladdar kolfångsten
Cyanobakteriella karboxysomer är små, ikosaedriska proteinkompartiment som ligger inne i cellen. Deras proteinskal släpper in vätekarbonatjoner samtidigt som de bromsar utflödet av CO2, vilket skapar en CO2‑rik bubbla runt Rubisco och dess partnerenzym karbanhydras (CA), som omvandlar vätekarbonat till CO2. Denna CO2‑koncentrationsmekanism låter cyanobakterier frodas även när CO2 i omgivningen är knapp. Eftersom den Rubisco‑form som används i dessa karboxysomer liknar växternas Rubisco, ses de av forskare som en realistisk modell för att omkonstruera växters kloroplaster för effektivare kolfångst.
Pusselbiten som talar med skalet
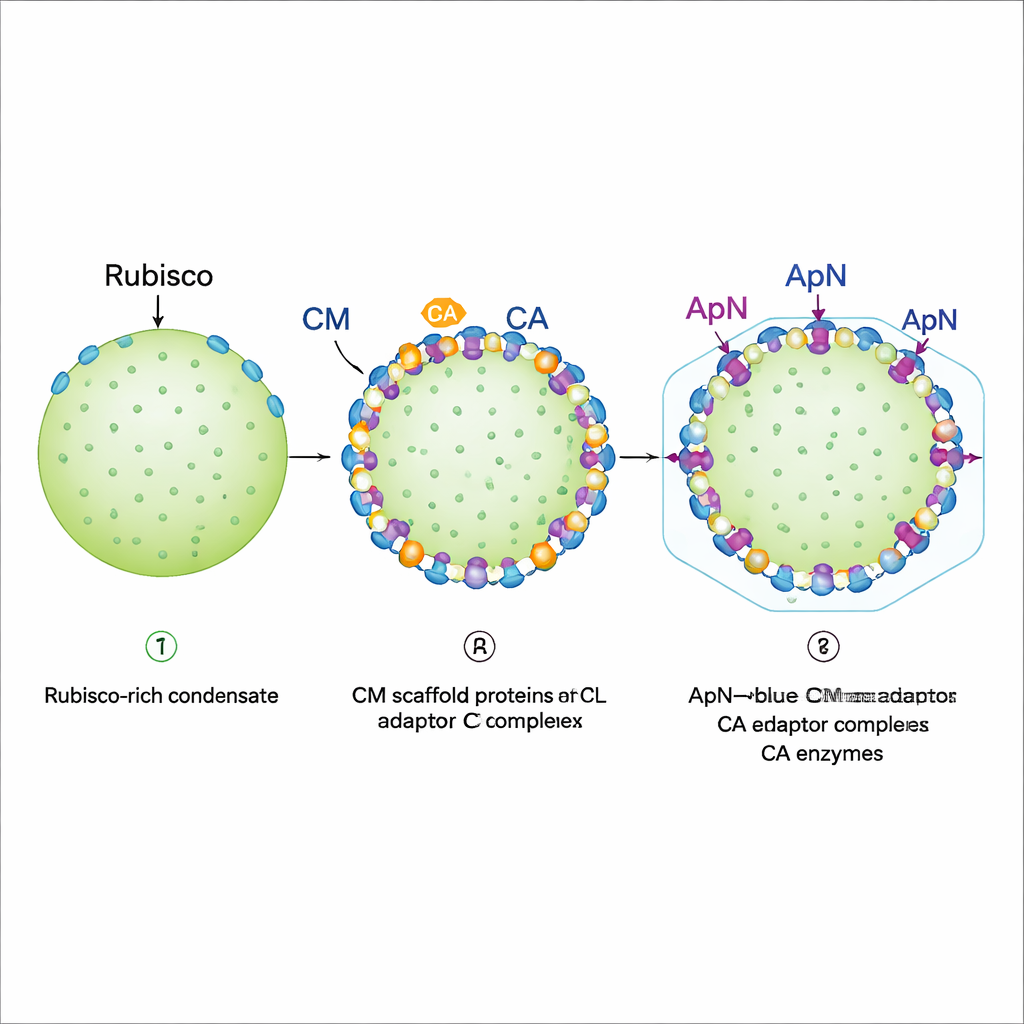
Innan det hårda proteinskalet formas bygger cyanobakterier först ett mjukt, droppliknande ”pro‑karboxysom”‑kondensat. Tidigare arbete visade att ett byggnadsproteinet CM samlar Rubisco och CA i denna droppe. Den saknade länken var hur skaladaptersproteinet ApN ansluter till denna struktur och senare kopplar den till skalproteinerna. Författarna fann att när ApN produceras ensamt bildar det en fyrdelad bunt (tetramer) som ignorerar de andra karboxysomkomponenterna. I detta tillstånd kan ApN inte dras in i droppar med Rubisco, CM och CA, vilket tyder på att en annan form av ApN måste användas under verklig karboxysom‑sammansättning.
Medbyggda adaptrar och en redox‑omkopplare
Genom att titta på den genetiska ordningen i cyanobakterier noterade teamet att apn ligger direkt efter cm i samma operon, så de två proteinerna brukar tillverkas tillsammans. Genom att efterlikna detta i bakterier samtidigt uttryckte de ApN och CM från en gemensam DNA‑sekvens. Under dessa förhållanden bildade ApN inte längre en fristående tetramer. Istället sammanfogades tre ApN‑subenheter och en CM‑subenhet till ett specifikt fyrdelat adapterkomplex, betecknat (ApN)3:CM. Detta hetero‑komplex fäste lätt vid Rubisco, CA och CM och kunde gå med i kondensat. Strukturell modellering och kryo‑elektronmikroskopi visade att ApN och CM möts via en finjusterad ”tunga‑och‑ränna”‑yta; när viktiga ApN‑rester i detta gränssnitt muterades bildades inte adaptern och cellerna misslyckades med att bygga korrekta karboxysomer.
Positionering i kanten och förberedelse för skalet
Fluorescensavbildning av rekonstituerade droppar avslöjade ett stegvis monteringsprogram. Först bildar Rubisco, fullängds‑CM och en kortare CM‑variant ett tätt kondensat som beter sig som en gel: Rubisco är i praktiken låst på plats. Därefter tillsätts CA och (ApN)3:CM‑komplexet. Bägge föredrar att lägga sig i den yttre kanten av den befintliga droppen, förutsatt att CA anländer före eller samtidigt som (ApN)3:CM. Denna kantlokalisering är avgörande eftersom ApN:s svans bär en ”inkapslingspeptid” som binder skalproteiner. Författarna upptäckte också en inbyggd redox‑omkopplare: när insidan av det mognande karboxysomet blir mer oxiderande kan två konserverade cysteinrester i ApN oxideras, vilket skjuter adaptern från en (ApN)3:CM‑tetramer mot en (ApN)2:CM‑trimer. I levande cyanobakterieceller gjorde förändringar av dessa cysteiner att tillväxten saktade och gav upphov till förstorade men fortfarande funktionella karboxysomer, vilket understryker deras roll i att finjustera mognaden.
Lärdomar för att bygga bättre grödor
För en icke‑specialist kan dessa molekylära detaljer låta torra, men de kokar ner till ett enkelt budskap: timing och partnerskap betyder något. Arbetet visar att ApN måste samproduceras med CM för att undvika att bilda en värdelös självklunga och istället bli en adapter som parkerar på ytan av pro‑karboxysomet, redo att rekrytera skalet. Ett redox‑känsligt steg hjälper sedan till att omforma denna adapter när kompartimentet sluts och dess interna kemi förändras. För växtingenjörer som hoppas överföra cyanobakteriell CO2‑koncentrationsmaskineri till grödor framhäver studien att det inte räcker att flytta enskilda delar; man måste också respektera ordning, kvoter och kemiska triggerfaktorer som styr karboxysom‑sammansättningen i naturen.
Citering: Zang, K., Hong, X., Nguyen, N.D. et al. Stages of biomolecular condensate formation in pro-β-carboxysome assembly. Nat. Plants 12, 447–464 (2026). https://doi.org/10.1038/s41477-026-02227-6
Nyckelord: karboxysom‑sammansättning, cyanobakteriell CO2‑koncentrationsmekanism, Rubisco‑kondensat, biomolekylära kondensat, syntetisk fotosyntes