Clear Sky Science · sv
Transkriptionell kompetens bestämmer den heterokromatin‑nukleerande potentialen hos isolerade MSR‑enheter
Dolda strömbrytare i vårt DNA
Våra genom är kompakta i cellkärnan i två huvudtillstånd: aktiva regioner som rymmer gener, och tätt packade sträckor som länge betraktats som genetiskt ”mörk materia”. Denna studie ställer en föga komplicerad fråga: vad får ett DNA‑stycke att bli detta tätt packade, gen‑tystande material—kallat heterokromatin—från början? Genom att dissekera en specifik klass av repetitivt DNA i mus visar författarna att inte alla repetitioner är likadana: endast de som kan stödja en särskild typ av transkription kan slå omkopplaren som bygger och upprätthåller dessa tysta DNA‑områden.
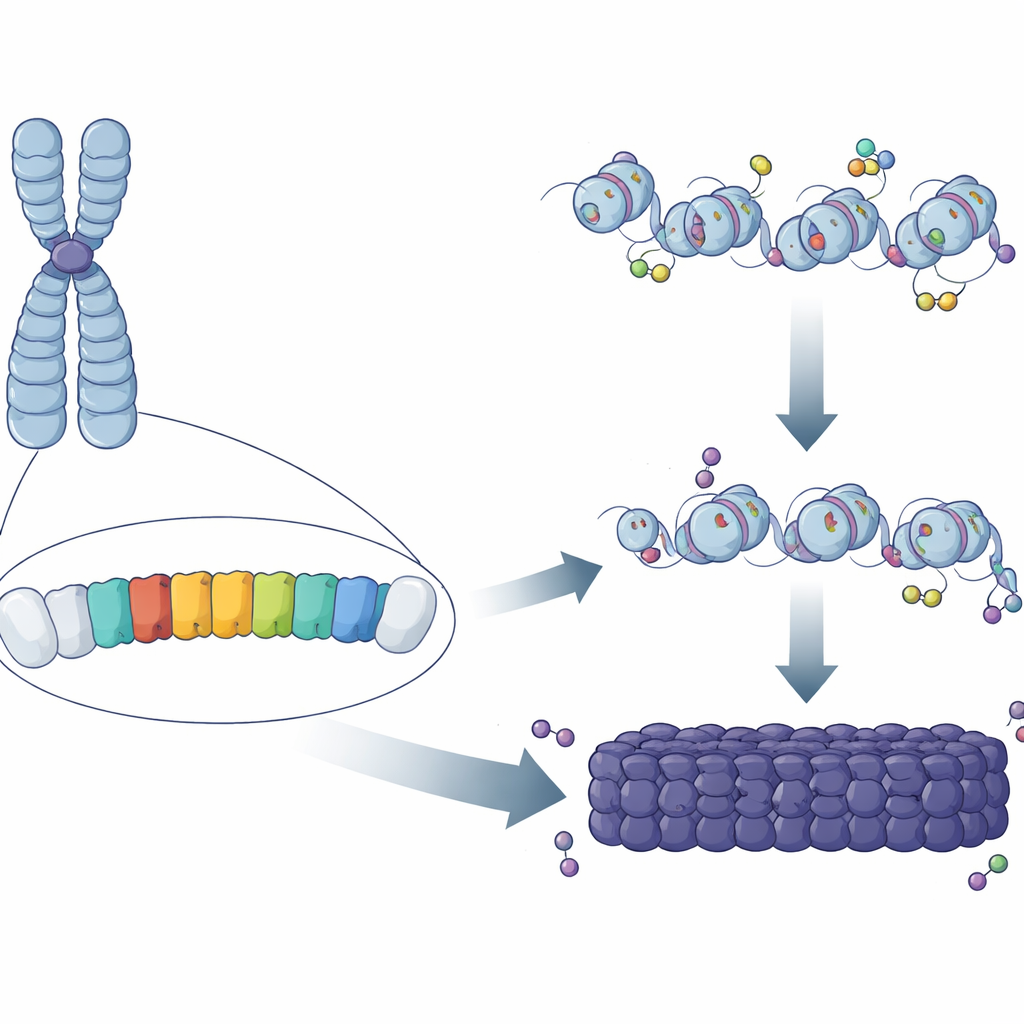
Repeterande mönster i genomet
Nästan hälften av däggdjurs DNA består av repeterade sekvenser, många klustrade i regioner kring kromosomens centrum. I mus är en huvudkomponent i dessa regioner ”major satellite repeat” (MSR), en kort A/T‑rik DNA‑sekvens kopierad hundratusentals gånger. Klassiskt arbete visade att dessa regioner täcks av kemiska märkningar och proteiner som låser DNA i ett kompakt, skyddande tillstånd. Men det var fortfarande oklart varför vissa MSR‑kopior blir fullständigt heterokromatiska medan andra, utspridda annorstädes i genomet, inte gör det. Författarna antog att små skillnader i sekvens eller beteende hos enskilda MSR‑enheter kan avgöra om de kan starta, eller ”nukleera”, ett område av heterokromatin.
Bygga en testplats i genomet
För att testa idén på ett rent sätt konstruerade teamet mus‑embryonala stamceller med en artificiell landningsplatta i ett tyst område på kromosom 2—en region utan närliggande gener eller repetitioner och utan detekterbar aktivitet. I denna neutrala plats infogade de olika DNA‑fragment: intakta MSR‑enheter, kraftigt omkastade MSR‑varianter, och kontrollelement såsom virala promotorer eller segment av mobila element. Detta gjorde det möjligt att enhet för enhet undersöka vilka sekvenser som kan attrahera de kännetecknande heterokromatin‑dragen: en specifik kemisk märkning på histonproteiner (H3K9me3), bindning av HP1‑proteiner, och införandet av länk‑histon H1—alla tillsammans som förtjockar och stabiliserar det lokala kromatinet.
Endast transkriptionsberedda repetitioner etablerar tyst kromatin
Resultaten var påfallande selektiva. En enda intakt MSR‑enhet infogad vid testplatsen var inte tillräcklig för att förändra kromatinet. Däremot konverterade tre eller fler tandade kopior av den intakta MSR‑sekvensen den omgivande regionen till en heterokromatin ”ö”, med stark H3K9me3, HP1 och histon H1. Till skillnad härifrån misslyckades lika långa sträckor av omkastad MSR‑sekvens, eller av en annan repetitionstyp (LINE‑1 5' oöversatta region), att göra det, även om de kunde driva stark transkription. Den avgörande skillnaden var att flerkopior av intakta MSR‑enheter stödde en måttlig, bidirelativ transkription som producerade korta, icke‑typiska RNA‑molekyler tätt associerade med kromatin. Detta mönster, snarare än hög, gen‑liknande transkription, korrelerade med förmågan att nukleera heterokromatin.
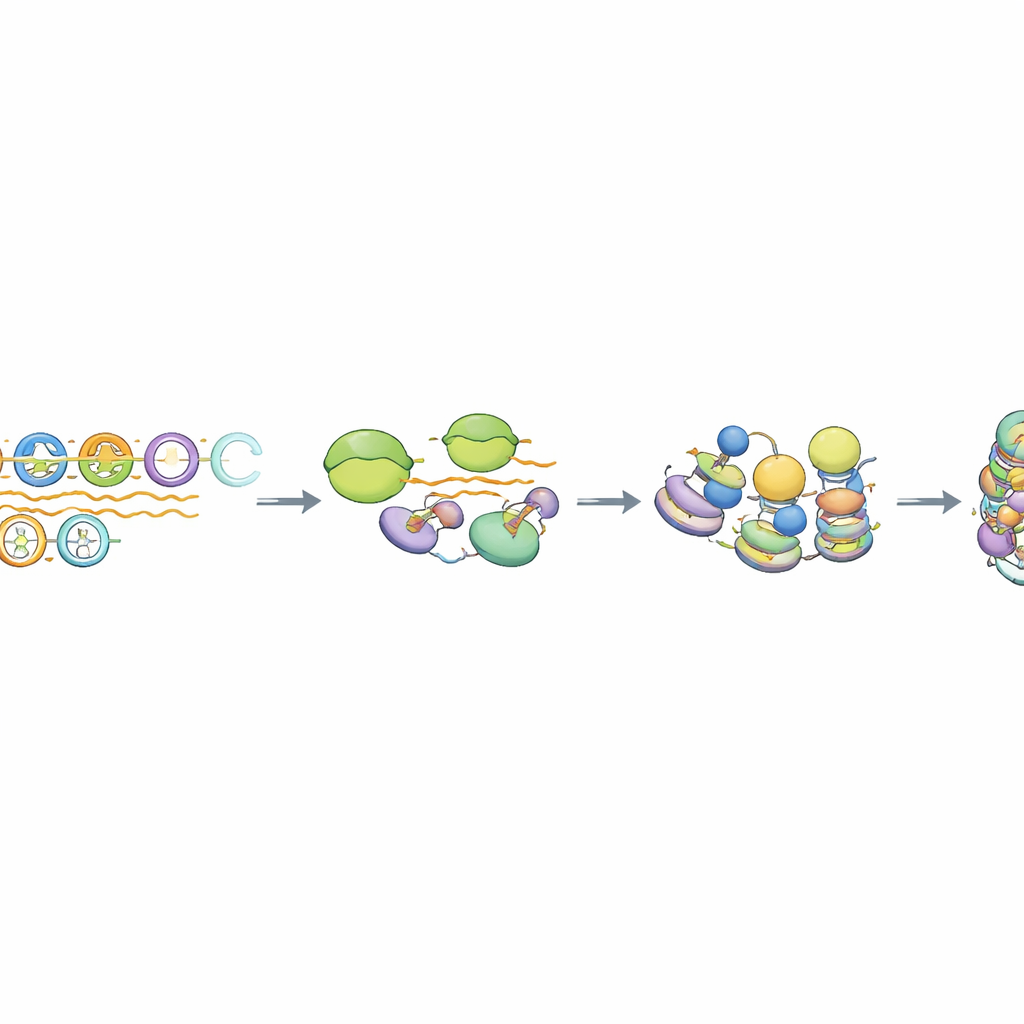
En särskild form av transkription och RNA‑bearbetning
Vid närmare granskning fann författarna att RNA‑polymeras II—enzymet som normalt syntetiserar budbärar‑RNA—kort engagerar MSR‑arrayer men inte effektivt förlänger till långa transkript. De resulterande RNA‑na saknar typiska mRNA‑kännetecken såsom en skyddande 5'‑lock och långa poly‑A‑svansar, och de förblir nära det DNA som producerade dem. Ett proteinkomplex kallat Integrator, känt för att trimma och terminera vissa icke‑kodande RNA, var specifikt förhöjt vid intakta MSR‑enheter. När Integratorns klyvningsaktivitet minskades steg nivåerna av MSR‑deriverade RNA markant, men den viktiga repressiva histonmarkeringen bestod medan HP1‑bindningen förändrades subtilt. Genom‑omfattande analys visade att endast de mest intakta ~10–15 % av MSR‑kopiorna beter sig på detta sätt, vilket framhäver en undergrupp av ”kompetenta” repetitioner kopplade till denna transkriptionsberoende tystnads‑väg.
Uppvridet DNA som promotorimitat
Teamet undersökte också hur MSR‑DNA självt uppmuntrar denna ovanliga transkription. Flerkopiorade MSR‑arrayer, men inte enkel‑ eller dubbla kopior, visade tydliga tecken på lokalt uppvridet DNA och RNA:DNA‑hybrider, strukturella egenskaper som ofta ses nära aktiva promotorer och på pausställen. Dessa konfigurationer förstärktes när topoisomeras‑enzymer hämmades, och de sammanföll med ökad MSR‑transkription och starkare heterokromatin‑drag. Författarna föreslår att tre eller fler tandemkopior av MSR skapar en fysisk DNA‑topologi som imiterar en promotor, vilket inbjuder polymeras och transkriptionsfaktorer att engagera sig precis tillräckligt för att generera korta RNA som, tillsammans med specifika proteiner, förstärker en kompakt kromatinarkitektur.
Varför detta betyder något för genomhälsan
För en lekman visar detta arbete att delar av vårt ”skräp”‑DNA fungerar som noggrant avstämda strömbrytare, som använder en blandning av DNA‑form, låg nivåtranskription och RNA‑bearbetning för att bygga genomets skyddande skikt. Endast MSR‑enheter som kan stödja denna kontrollerade, icke‑budbärar‑transkription kan tända ny heterokromatin, medan omkastade eller alltför aktiva element inte kan det. Denna DNA/RNA‑baserade logik hjälper till att förklara hur celler skiljer mellan regioner som ska hållas tysta och dem som får hysa gener, och varför felreglerade satellit‑RNA är kopplade till cancer och utvecklingsstörningar. I grunden visar studien att genomets repetitiva ”bakgrund” inte är passivt fyllnadsmaterial utan en aktiv ingenjör av nukleär arkitektur och stabilitet.
Citering: Lo, YH., Shukeir, N., Erikson, G. et al. Transcriptional competence defines the heterochromatin nucleating potential of isolated MSR units. Nat Commun 17, 2653 (2026). https://doi.org/10.1038/s41467-026-70991-2
Nyckelord: heterokromatin, satellit‑DNA, icke‑kodande RNA, kromatinstruktur, genomstabilitet