Clear Sky Science · ru
Перфорантный путь и афференты CA3–Шаффера согласованно регулируют пространственное обучение
Почему умение ориентироваться имеет значение
Будь то мышь, ищущая скрытую платформу, или человек, осваивающий новый город, мозг постоянно превращает зрительные, звуковые сигналы и информацию о собственном движении во внутреннюю карту. В этом исследовании изучается, как две ключевые области мозга — энторинальная кора и гиппокамп — совместно обеспечивают эту способность. Наблюдая в реальном времени активность тонких нервных волокон и точно стимулируя или подавляя отдельные пути у мышей, исследователи выяснили, как эти области координируют работу для формирования и стабилизации пространственных воспоминаний — выводы, которые в перспективе помогут понять и лечить расстройства памяти.
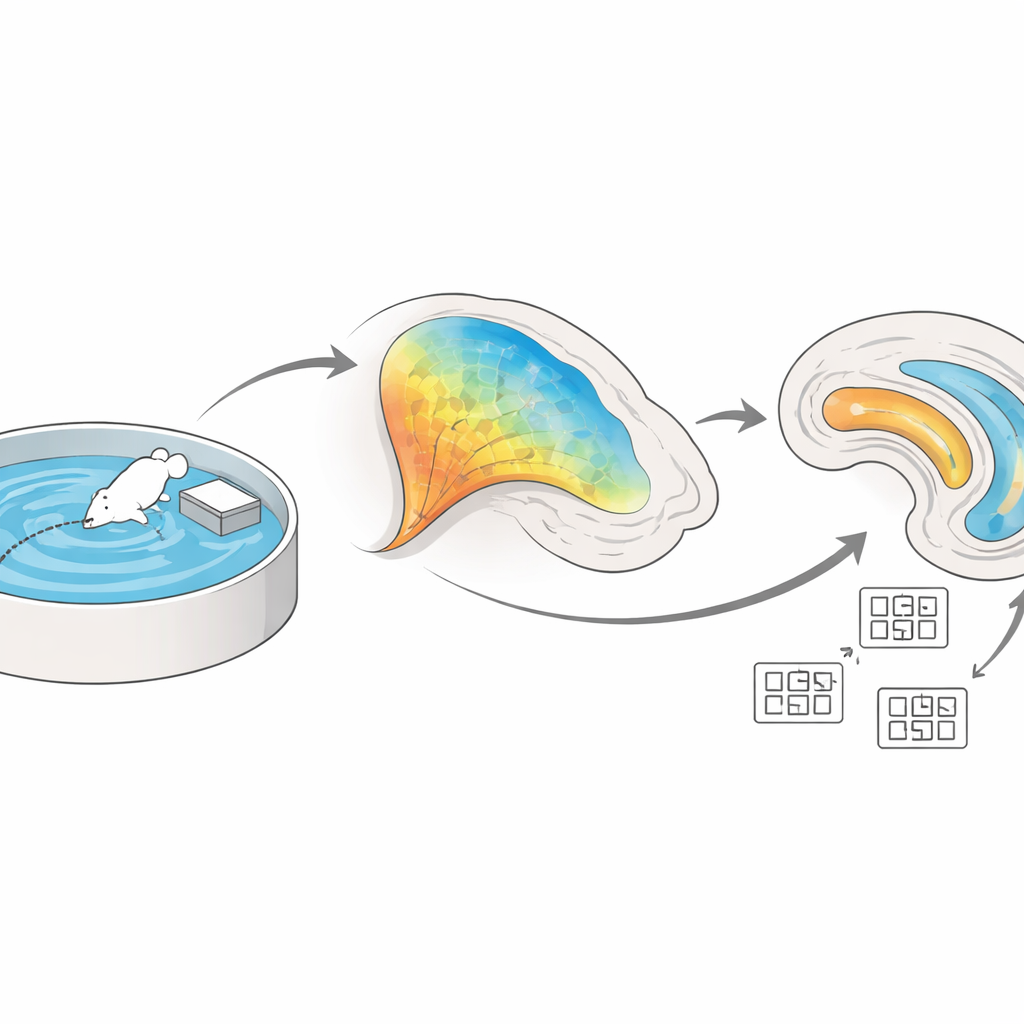
Два ключевых узла внутренней «GPS» мозга
Гиппокамп и прилегающая энторинальная кора расположены глубоко во внутренней части височной доли и формируют ядро нашей навигационной системы. Внутри гиппокампа существует внутренний контур, связывающий область CA3 с областью CA1 через волокна, известные как коллатерали Шаффера. Одновременно энторинальная кора посылает отдельный поток информации прямо в CA1 по так называемому перфорантному пути. Авторы поставили задачу выяснить, как эти два входных потока взаимодействуют при изучении животным планировки пространства и как изменения на их синапсах — в широком смысле «пластичность» — поддерживают построение устойчивой пространственной карты.
Наблюдая обучение по волокну
Чтобы проследить этот процесс в ведённых животных, команда использовала фиброзную фотореферометрию, технику, которая показывает активность нейронов через вспышки флуоресцентного света. Они генетически модифицировали мышей так, чтобы нейроны CA3, проецирующие в CA1, светились при активности, затем обучали животных в водном лабиринте Морриса — классическом тесте, в котором мышь должна запомнить скрытое положение небольшой платформы в бассейне. В ранних пробах волокна CA3–CA1 были сильно активны, пока животные искали; по мере того как мыши становились умелее и быстрее находили платформу, эта активность постепенно снижалась. Такая картина указывает на то, что эти связи особенно задействованы на этапе первоначального кодирования планировки окружения, а затем переходят в более эффективное, устойчивое представление после установления памяти.
Как вход сверху усиливает или притупляет обучение
Далее ученые спросили, как сигналы из энторинальной коры влияют на этот гиппокампальный контур. С помощью светочувствительных белков для активации энторинальных нейронов при записи активности волокон CA3–CA1 они показали, что стимуляция медиальной энторинальной коры надежно увеличивает активность этих гиппокампальных связей. Наоборот, когда они химогенно ослабляли сигналы энторинальной коры, достигающие CA1 во время обучения в водном лабиринте, активность CA3–CA1 уменьшалась, и мыши учились местоположению платформы медленнее и менее точно. В дополнительном наборе экспериментов авторы напрямую регистрировали активность энторинальных нейронов, проецирующих в CA1, и наблюдали, что их активность нарастала в течение дней тренировки, отражая улучшение поведения животных. В совокупности эти результаты указывают, что сильные, хорошо скоординированные сигналы энторинальной коры необходимы для правильной настройки гиппокампальных контуров для навигации.
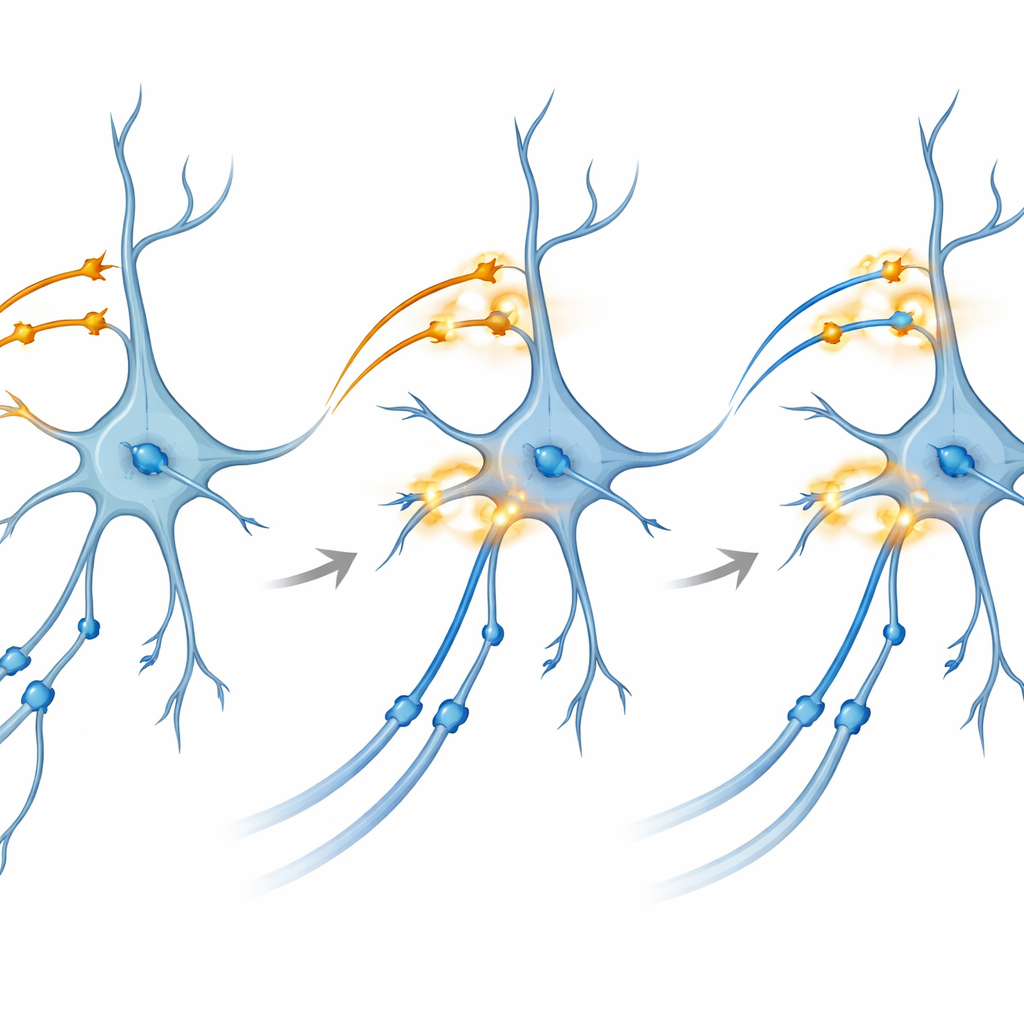
Приближение к тому, как укрепляются связи
Чтобы исследовать базовый механизм, команда перешла к живым срезам мозга, сохраняя их жизнеспособность в чашке Петри. Здесь они могли раздельно управлять светочувствительными белками в CA3 и в энторинальных входах в CA1. Удивительно, но тета‑всплесковая световая стимуляция — паттерны, имитирующие естественную ритмическую разрядку — примененная к каждому пути по отдельности, не вызывала надежного длительного потенцирования, длительного усиления синапсов, которое принято связывать с памятью. Однако при подаче точно синхронизированного двухцветного всплеска, коактивирующего оба пути в CA1 одновременно, связи CA3–CA1 демонстрировали устойчивое и продолжительное усиление. Блокировка NMDA‑рецепторов или специфических кальциевых каналов, двух хорошо известных молекулярных ворот пластичности, предотвращала этот эффект, связывая явление с классическими биохимическими путями формирования памяти.
Что это значит для памяти и болезней
В целом исследование рисует картину пространственного обучения как скоординированного танца между входящими сигналами энторинальной коры и внутренней проводкой гиппокампа. Энторинальные входы не только несут информацию о положении и контексте; они также действуют как мощный регулятор, который может усилить или ослабить активность в волокнах CA3–CA1 и, при коактивации, вызвать длительное укрепление этих синапсов. Для непрофессионального читателя ключевой вывод состоит в том, что «GPS» мозга не опирается на одну область, а на партнерство, в котором одна структура задает сцену, а другая закрепляет детали. Нарушения этого партнерства — будь то из‑за старения, травмы или нейродегенеративных заболеваний — могут объяснять, почему люди теряются в знакомых местах, и нацеливание на эти пути может открыть новые возможности для сохранения или восстановления пространственной памяти.
Цитирование: Huang, F., Temitayo Bello, S., Lau, S.H. et al. The perforant pathway and CA3-Schaffer collateral afferents coordinate to regulate spatial learning. Commun Biol 9, 364 (2026). https://doi.org/10.1038/s42003-026-09577-z
Ключевые слова: пространственная память, гиппокамп, энторинальная кора, синаптическая пластичность, навигация