Clear Sky Science · ru
Белок-адаптор субстрата Cullin 3 1 (MtCSP1) модулирует образование узелков через взаимодействие с GTPазой ARFA1
Как бобы производят собственное удобрение
Современное сельское хозяйство сильно зависит от азотных удобрений, которые повышают урожайность, но наносят вред окружающей среде. Бобовые растения — такие как клевер, горох и люцерна — предлагают естественную альтернативу: они размещают дружелюбные почвенные бактерии в специализированных корневых структурах, называемых узелками, где атмосферный азот превращается в питательные вещества для растения. В этом исследовании показано, как ранее неохарактеризованный растительный белок MtCSP1 помогает контролировать формирование этих узелков, тонко регулируя внутриклеточный трафик и утилизацию ключевых молекулярных переключателей в клетках корня.
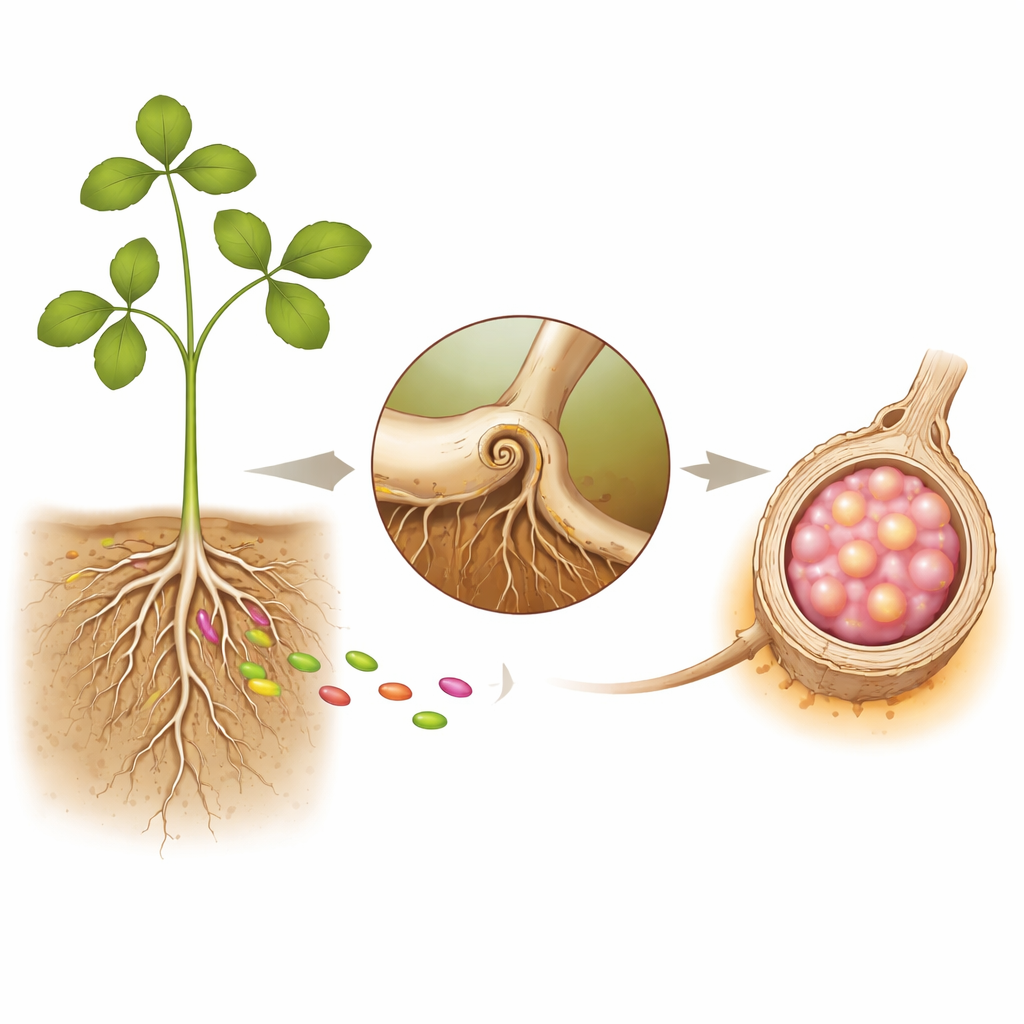
Корни растений и их бактериальные жильцы
Бобовые вступают в партнёрство с ризобиями — почвенными бактериями, способными переводить атмосферный азот в доступные формы, например аммоний. Чтобы безопасно разместить этих бактерий, корни формируют узелки — крошечные «фабрики» на поверхности корня. Процесс начинается, когда бактерии прикрепляются к корневым волоскам, которые скручиваются и образуют узкий канал, называемый ниткой инфекции. Через этот канал бактерии продвигаются в более глубокие ткани корня, где начинает развиваться новый орган — узелок. В зрелом узелке бактерии окружены растительными мембранами и превращаются в специализированные «бактероиды», которые фиксируют азот для растения в обмен на сахара и энергию.
Клеточные регулировщики трафика в корневых клетках
Построение нитей инфекции и узелков требует интенсивной перестройки мембран растительных клеток. Эта перестройка зависит от везикул — небольших мембранных пузырьков, которые переносят вещества внутри клетки. Их перемещение и время активности контролируются малыми молекулярными переключателями, известными как GTPазы. Один из таких переключателей, ARFA1, помогает формировать и направлять везикулы у многих организмов. Но в контексте узелков, фиксирующих азот, было неясно, как регулируется ARFA1 и как долго он остаётся активным до утилизации.
Новая связь между трафиком и переработкой
Исследователи искали растительные белки, которые физически взаимодействуют с ARFA1 в модельной бобовой Medicago truncatula. С помощью системы двухгибридного анализа в дрожжах и тестов взаимодействия белков в листьях растений они идентифицировали MtCSP1 — белок, который относится к семейству адаптеров, связывающих конкретные мишени с клеточной «измельчительной» системой убиквитина. MtCSP1 содержит домен BTB/POZ, характерный для белков, привлекающих выбранных партнёров к комплексам на основе Cullin 3, которые отмечают белки для разрушения. Флуоресцентная визуализация показала, что MtCSP1 и ARFA1 встречаются на везикулах, расположенных в поздних эндосомах — клеточных перевалочных пунктах, которые часто направляют груз на деградацию.
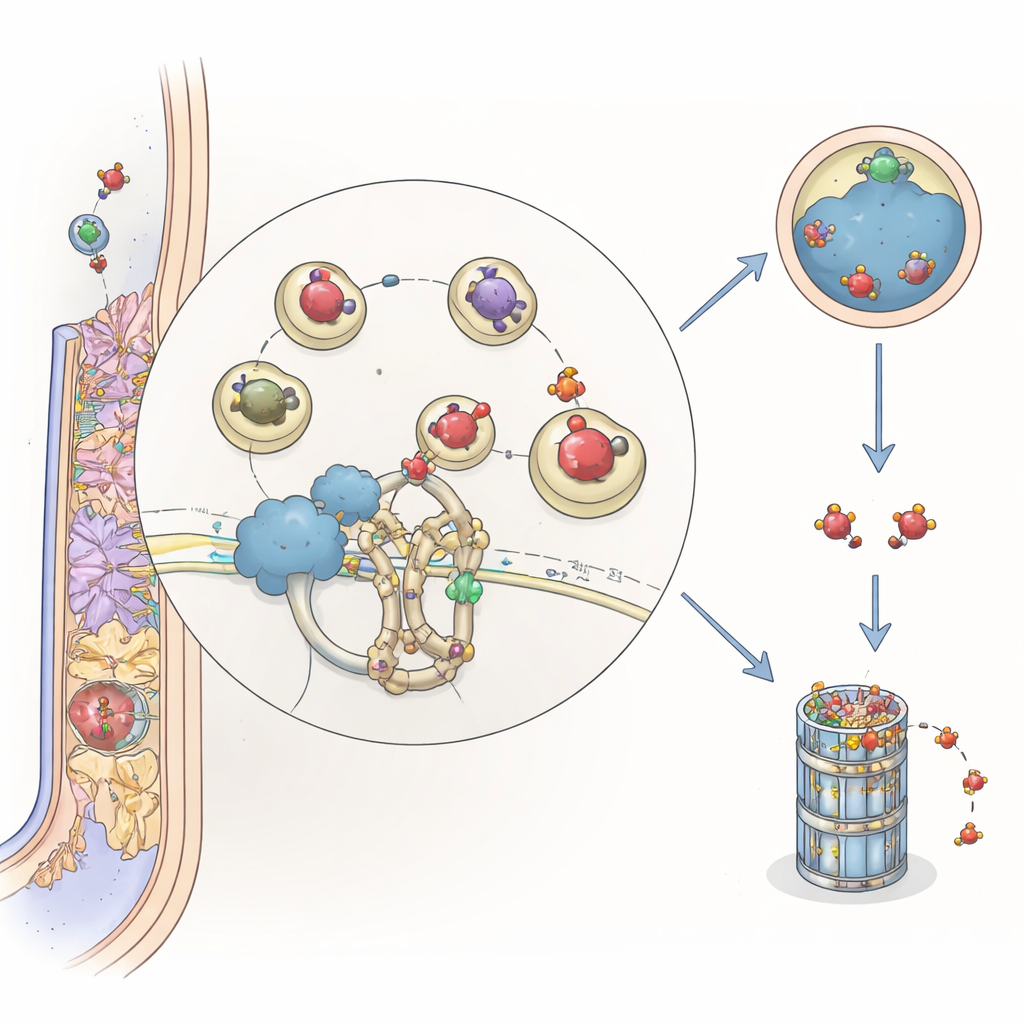
Когда и где действует новый игрок
Чтобы понять, когда используется MtCSP1, команда отслеживала активность его промотора — участока ДНК, контролирующего синтез белка. Они обнаружили, что MtCSP1 включается в кончиках корней, в появляющихся боковых корнях и в ключевых областях развивающихся и зрелых узелков, особенно в меристеме (зоне роста) и инфекционных зонах. Общедоступные наборы данных по экспрессии генов показали, что MtCSP1 и ARFA1 часто активируются совместно, что указывает на координацию присутствия переключателя (ARFA1) и адаптера (MtCSP1) растением во время формирования корневых органов.
Тонкая настройка числа узелков и хода инфекции
Затем учёные изменили уровни MtCSP1 в корнях. При подавлении MtCSP1 методом РНК-интерференции растения со временем образовывали меньше узелков, и многие нитки инфекции застревали в корневых волосках, не достигая внутренних слоёв корня. Тем не менее образовавшиеся узелки были нормального размера, что указывает на то, что MtCSP1 в большей степени влияет на инициацию и продвижение инфекции, а не на последующий рост. Напротив, при сверхэкспрессии MtCSP1 корни развивали больше узелков и демонстрировали изменения в прогрессии инфекционных событий, вновь без существенных изменений размера или формы узелков. Эти результаты свидетельствуют о том, что MtCSP1 не обязателен для запуска нитей инфекции, но критически важен для их правильного продвижения и успешного начала новых узелков.
Переработка переключателей для контроля симбиоза
Сопоставляя данные, авторы выдвигают гипотезу, что MtCSP1 действует как проводник, направляющий ARFA1, в активной или неактивной форме, к убиквитиновой машине Cullin 3 на везикулах поздних эндосом. Там ARFA1 может быть помечен и направлен либо в вакуоль для разрушения, либо в протеасому, что помогает выключать или корректировать везикулярный трафик после выполнения им своей функции. Регулируя продолжительность доступности ARFA1, MtCSP1 позволяет растению тонко настраивать деликатную хореографию нитей инфекции и образования узелков. Для непрофессионала смысл в том, что бобовые используют внутреннюю систему переработки, чтобы контролировать крошечные молекулярные светофоры внутри своих корней — обеспечивая эффективное и своевременное строительство своих природных «удобрительных фабрик».
Цитирование: Rípodas, C., Cretton, M., Eylenstein, A. et al. Cullin 3 substrate-adaptor protein 1 (MtCSP1) modulates nodulation through interaction with the GTPase ARFA1. Sci Rep 16, 8938 (2026). https://doi.org/10.1038/s41598-026-41112-2
Ключевые слова: фиксация азота, узелки бобовых, деградация белков, транспорт везикул, растительно‑микробный симбиоз