Clear Sky Science · ru
Функциональный переход пирамидных нейронов CA2 вдоль проксимодистальной оси определяет предпочтительную резонансную частоту
Как мозговые ритмы формируют память и социальное поведение
Гиппокамп — глубокая структура мозга, важная для формирования воспоминаний, ориентирования в пространстве и управления социальным поведением. Внутри него лежит узкая, но влиятельная область CA2, долгое время оставшаяся в тени своих более известных соседей CA1 и CA3. В этом исследовании поставлен, на первый взгляд, простой, но далеко идущий вопрос: «слушают» ли нервные клетки CA2 вдоль этой тонкой полоски ткани разные мозговые ритмы, и может ли это помочь объяснить, как гиппокамп координирует сложные мысли и поведение? 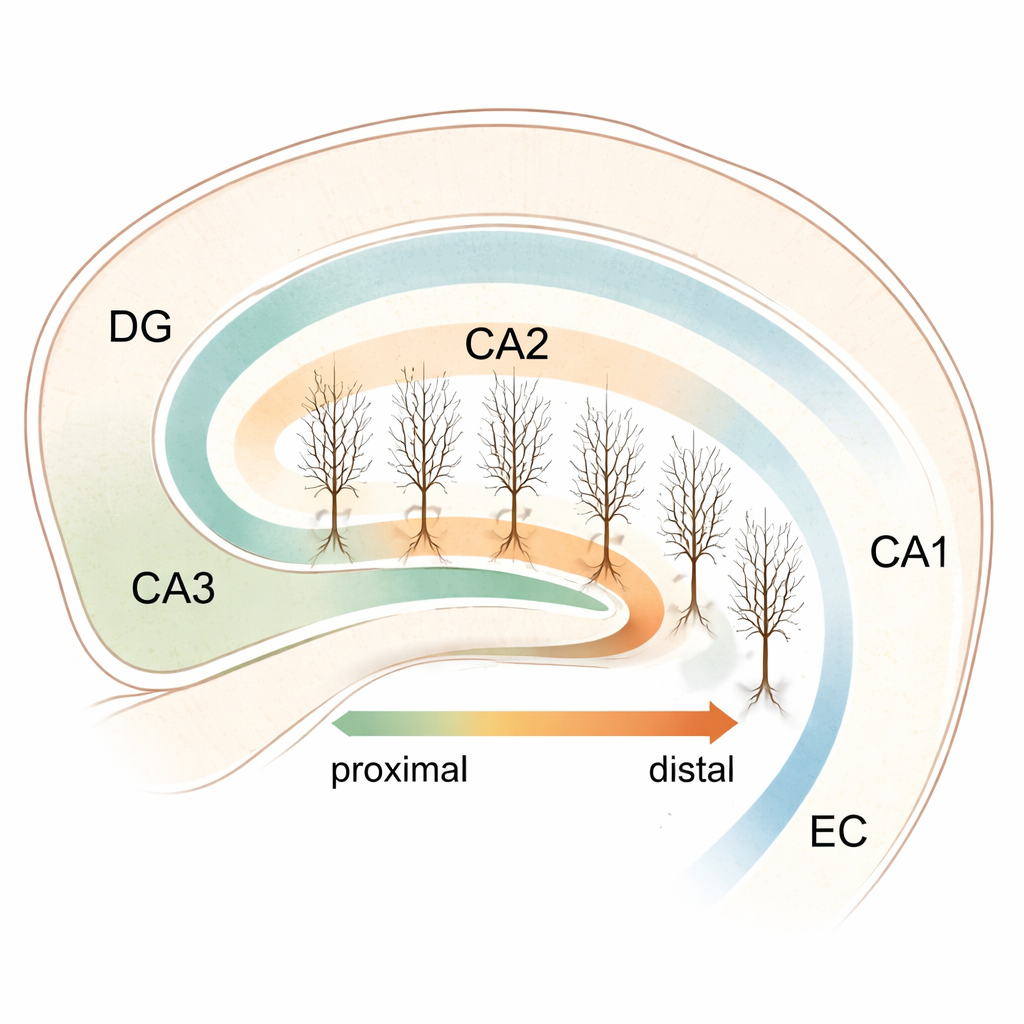
Скрытая область в цепи памяти
CA2 располагается между двумя крупными узлами гиппокампа — CA3, который способствует генерации быстрых сетевых всплесков, и CA1, тесно связанным с более медленными, ритмичными волнами, возникающими во время движения и обработки памяти. CA2 связывали с социальной памятью, регуляцией агрессии и пространственной ориентацией, однако его внутренняя организация оставалась неясной. Чтобы исследовать это, ученые выращивали тонкие срезы гиппокампа мыши вместе с энторинальной корой — важной областью входа — в длительной культуре. Такой подход сохраняет большую часть естественной проводящей архитектуры и при этом избегает повреждений от резки свежей ткани, что позволило изучать отдельные клетки CA2 в стабильных условиях.
Одинаковая форма, разные внутренние настройки
С помощью молекулярного маркера PCP4 команда точно очертила область CA2 и присвоила каждой пирамидной клетке «проксимодистальную» позицию: ближе к границе с CA3 (проксимально) или ближе к CA1 (дистально), с промежуточными значениями вдоль непрерывной оси. Затем они заполнили и реконструировали отдельные клетки в 3D, чтобы сравнить их ветвление. Несмотря на предыдущие намеки на вариации структуры CA2, исследователи не обнаружили заметной связи между положением клетки и ее общей дендритной формой — число ветвей, общая длина и точки разветвления оставались в целом схожими по всей полоске. Параллельно они измеряли спонтанные возбуждающие сигналы, поступающие к этим нейронам, и снова не увидели четкого градиента: базовый размер и частота входящих синаптических событий были относительно однородны от одного конца CA2 до другого. Это указывало на то, что если CA2 функционально разделен, ключевые различия могут заключаться во внутренних электрических настройках, а не в самой проводящей схеме или силе входов.
Постепенный сдвиг электрической возбудимости
Когда исследователи вводили токи непосредственно в клетки CA2 и отслеживали реакцию их мембран, проявились ясные позиционные тенденции. Клетки, близкие к CA3, имели более высокое входное сопротивление, то есть небольшие токи вызывали большие изменения напряжения, и они были менее склонны к «сагу» — характерному отскоку, наблюдаемому при кратковременном гиперполяризующем воздействии. По мере продвижения к CA1 входное сопротивление снижалось, тогда как саг и связанный с ним отскок становились более выраженными. Также системно менялись потенциалы действия: дистальные клетки требовали меньшего тока для генерации спайка, легче реагировали на одинаковый уровень входного возбуждения и демонстрировали тонкие сдвиги в форме спайка. Иными словами, нейроны CA2 обладают в целом схожей анатомией, но функционируют с постепенно настроенными электрическими параметрами вдоль проксимодистальной оси, что делает одни более возбудимыми и динамично отзывчивыми, чем другие.
Настройка на разные диапазоны мозговых волн
Одно из самых впечатляющих открытий касалось того, как эти клетки реагируют на ритмические входы на разных частотах — свойства, известного как субпороговая резонанс. Подавая на мембрану мягкие синусоидальные токи, меняя частоту от медленных к более быстрым циклам, команда могла определить, на какой частоте колебания напряжения клетки усиливаются наиболее заметно. Проксимальные нейроны CA2 проявляли мало или вовсе не проявляли предпочтений; они вели себя как универсалы. Дистальные же клетки все более четко демонстрировали резонансные пики, которые смещались от очень медленного дельта-диапазона в сторону нижней теты — порядка нескольких циклов в секунду. Поскольку тета-ритмы доминируют во время исследования окружающей среды, навигации и кодирования памяти, этот градиент указывает на то, что дистальные клетки CA2 естественно настроены захватывать эти поведенческие ритмы, вероятно, за счет градуированной активности ионных каналов, которые также порождают саг. 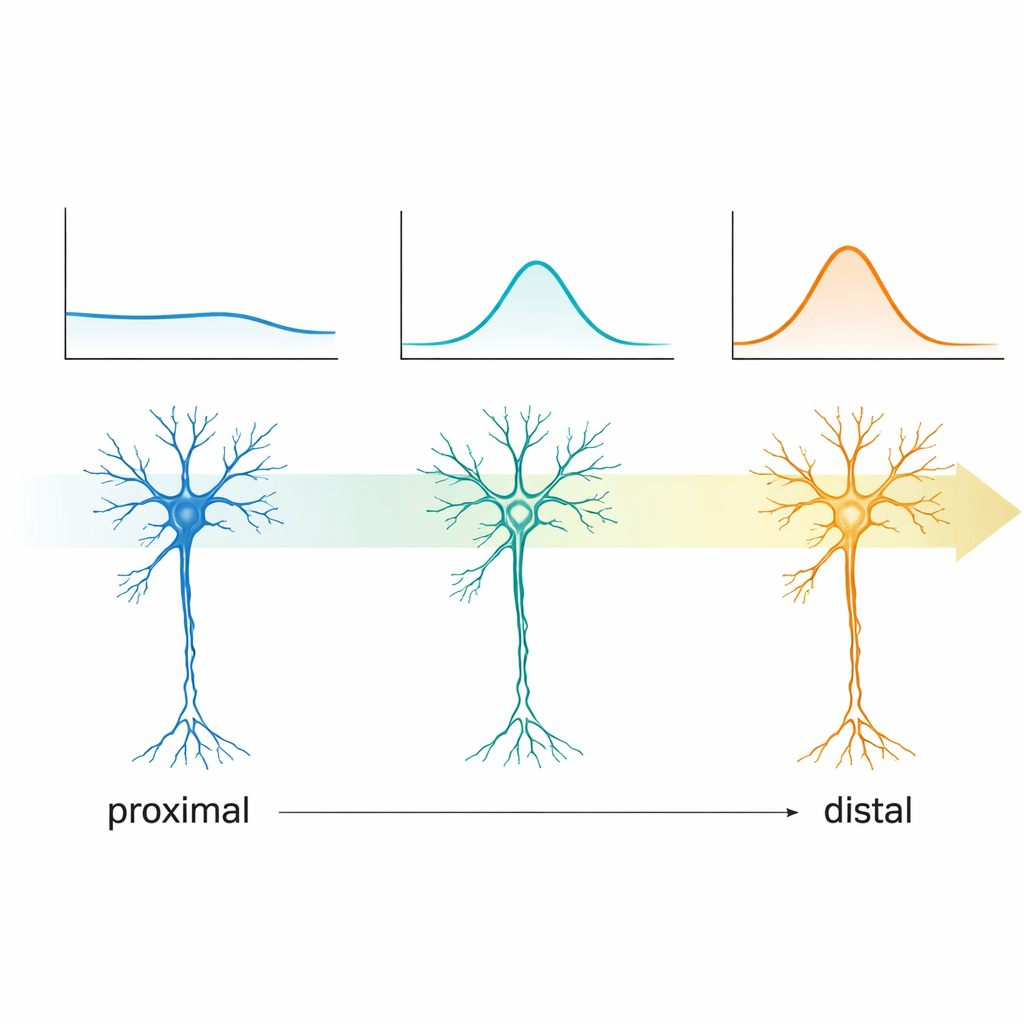
Тонкий градиент с большими сетевыми последствиями
В сумме работа показывает, что CA2 — это не однородная полоска идентичных нейронов, а плавная переходная зона между двумя разными режимами работы гиппокампа. Ближе к CA3 клетки CA2 лишены выраженной резонансности и, возможно, лучше подходят для участия в кратких, быстрых событиях, таких как sharp-wave ripples, которые помогают проигрывать воспоминания во время отдыха и сна. В сторону CA1 клетки становятся более отзывчивыми на тета-ритмы и, вероятно, предпочтительно связаны с энторинальными входами, несущими информацию о положении, контексте и текущем опыте. Для неспециалиста главный вывод таков: даже на крошечных расстояниях в мозге нейроны могут быть тонко настроены на разные «станции» шкалы мозговых волн, что позволяет небольшой области вроде CA2 гибко перенаправлять и формировать информацию, лежащую в основе памяти, навигации и социального поведения.
Цитирование: Kruse, P., Eichler, A., Brockmeyer, K. et al. Functional transition of CA2 pyramidal neurons along the proximodistal axis determines resonance frequency preference. Sci Rep 16, 7172 (2026). https://doi.org/10.1038/s41598-026-39754-3
Ключевые слова: гиппокамп, нейроны CA2, осцилляции мозга, тета-ритм, цепи памяти