Clear Sky Science · ru
Связь гиппокампа и супрамаммиллярной области во время сна и бодрствования
Почему это важно для повседневной памяти
Каждую ночь, во сне и даже в тихом покое, наш мозг воспроизводит переживания и перестраивает воспоминания. В этом исследовании подробно рассматривается, как ключевой центр памяти — гиппокамп — «общается» с более глубокими структурами мозга, которые контролируют бодрствование, движение и настроение. Наблюдая за диалогом этих областей в разные состояния сна и бодрствования у крыс, исследователи показывают, как мозг гибко маршрутизирует информацию — порой посылая сильные направленные сигналы сверху вниз из цепочек памяти, а иногда позволяя глубоким структурам ненадолго взять на себя ведущую роль. Понимание этого обмена помогает объяснить, как сон поддерживает память, одновременно сохраняя стабильность базовых функций организма.
Двусторонние разговоры между центрами памяти и бодрствования
Команда записывала мелкие электрические сигналы из гиппокампа и двух связанных областей — супрамаммиллярного ядра и латеральной септы — у свободно передвигающихся крыс. Они отслеживали животных в состояниях тихого покоя, глубокого сна без быстрого движения глаз (NREM), сна с быстрым движением глаз (REM), богатого сновидениями, и во время активного бодрствования. На основании этих записей фиксировали кратковременные всплески синхронизированной активности гиппокампа, называемые рипплами, и связанные события в соседней области — зубчатой извилине (dentate gyrus). Одновременно измеряли частоту спайкинга нейронов в супрамаммиллярном ядре, которое влияет на бодрствование и стадии сна, и в латеральной септе, связывающей структуры памяти с системами мотивации и движения.
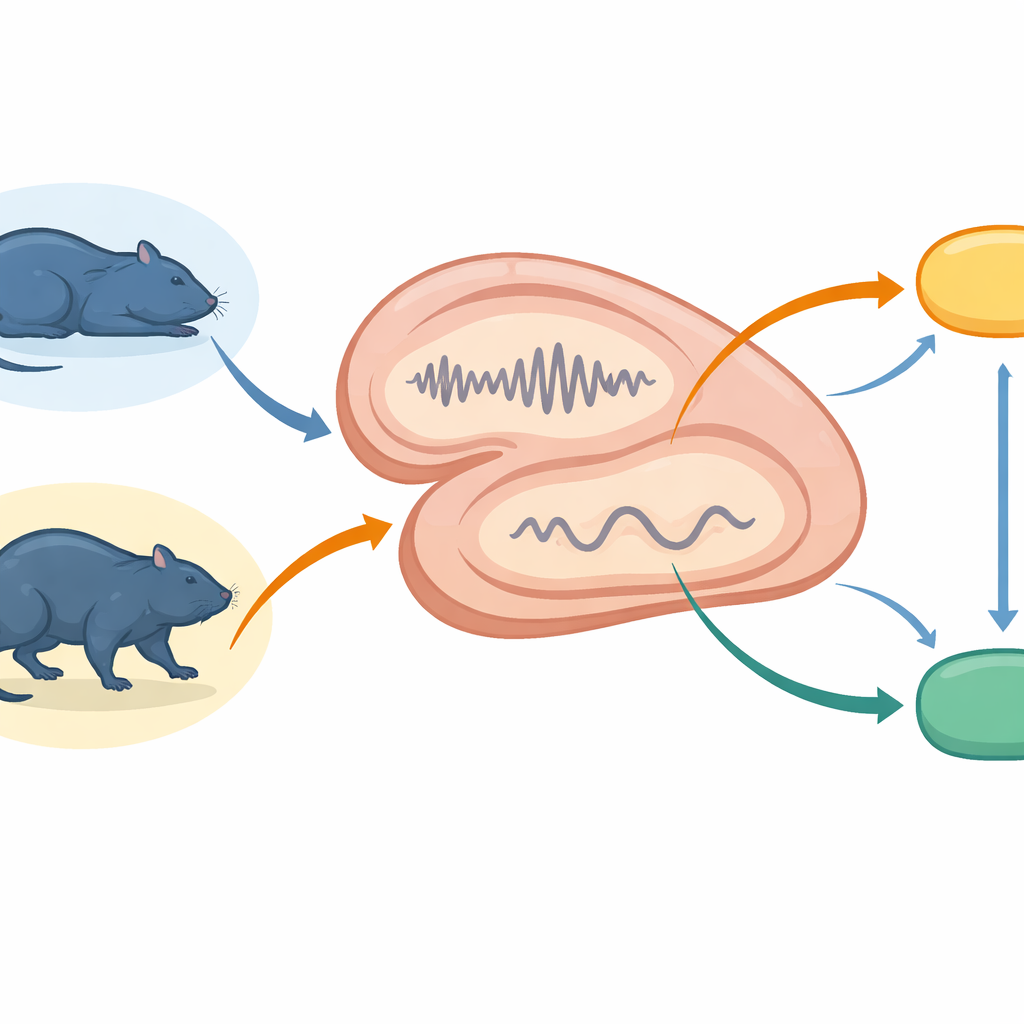
Сверху вниз — всплески в тихие моменты
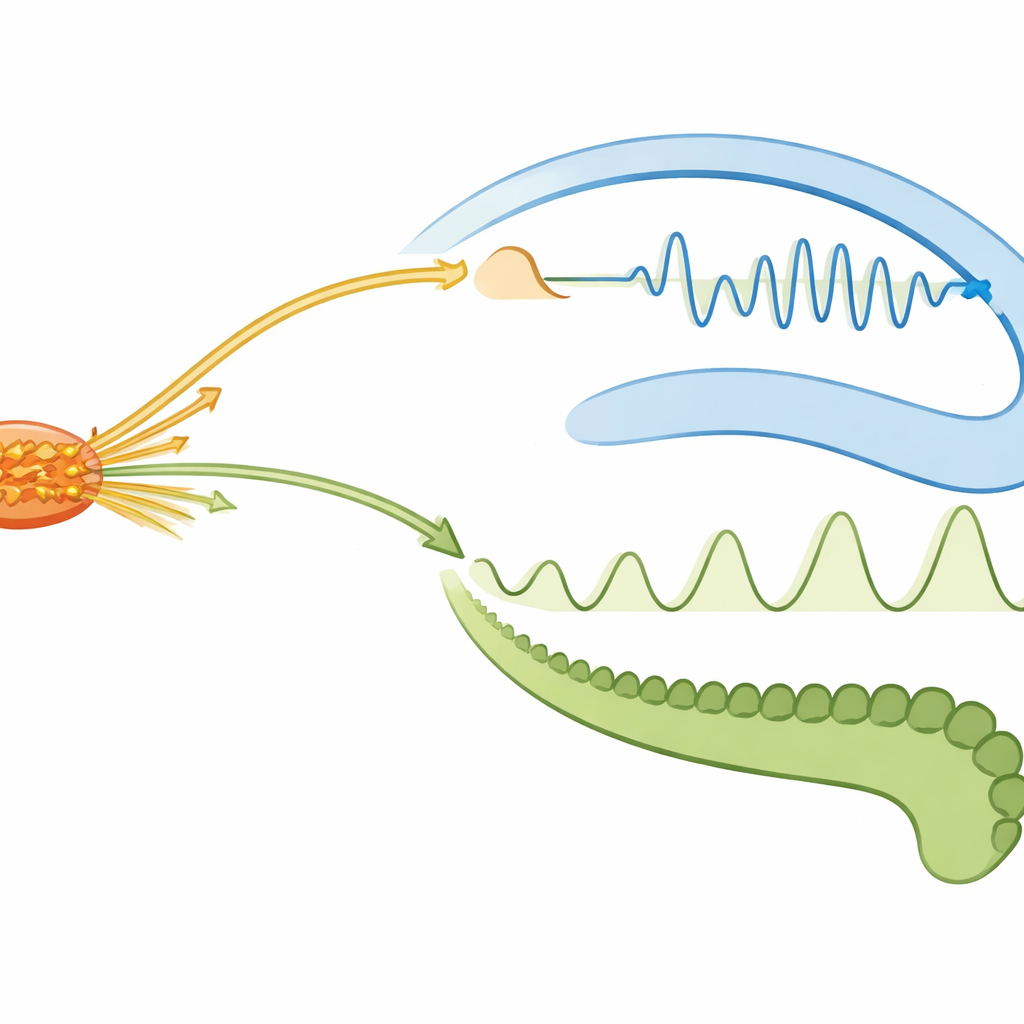
В периоды тихого бодрствования и NREM-сна — состояниях, предполагаемых благоприятными для реплея памяти — гиппокамп часто генерировал острые быстрые рипплы. Эти рипплы вызывали сильные локальные всплески активности внутри гиппокампа и более слабую, но надежную активацию в супрамаммиллярном ядре и латеральной септе. Проще говоря, система памяти рассылала краткие сигналы сверху вниз к более глубоким центрам, но с меньшей силой, чем внутри себя. Зубчатые спайки (dentate spikes), другой тип быстрых событий в соседнем субрегионе гиппокампа, вызывали более слабые и однородные ответы, которые едва достигали подкорковых участков. Это различие указывает на то, что рипплы — основной механизм быстрой и широкой координации, тогда как зубчатые спайки тонко настраивают локальную обработку, не оказывая сильного влияния на нисходящие центры.
Снизу вверх — толчки от глубоких вспышек
Коммуникация шла не в одну сторону. Когда супрамаммиллярное ядро входило в короткие периоды высокой активности, оно посылало сигналы снизу вверх обратно в гиппокамп. Их последствия сильно зависели от состояния мозга. Во время NREM-сна такие всплески кратковременно подавляли активность ключевой выходной области гиппокампа (CA1), тогда как нейроны зубчатой извилины реагировали более медленным усилением. В тихом бодрствовании те же супрамаммиллярные всплески вызывали менее выраженное падение активности CA1, но более четкое суммарное увеличение активности в гиппокампе. Эти закономерности показывают, что глубокие всплески могут либо ослаблять, либо усиливать выход гиппокампа, подстраивая, насколько сильно цепочки памяти влияют на остальную часть мозга, не отключая их полностью.
Ритмическая синхронизация в активных и сновидческих состояниях
Когда мозг переходил в состояния, богатые тета-ритмом — активное исследование в бодрствовании и яркие сновидения в REM — диалог менялся вновь. Тета-волны — более медленные ритмы, помогающие координировать спайкинг между областями. В активном бодрствовании тета организовывала повторяющийся порядок выстрелов: нейроны зубчатой извилины обычно активировались первыми в каждом цикле, затем латеральная септа, а затем CA1 около впадины волны, тогда как супрамаммиллярное ядро показывало мало согласованного временного сдвига. В этом состоянии супрамаммиллярные всплески вызывали быстрое, кратковременное ингибирование в CA1, но практически не затрагивали активность зубчатой извилины, что указывает на избирательную «затворку» некоторых входов гиппокампа. В REM-сне картина менялась: супрамаммиллярные нейроны теперь прочно синхронизировались с тета-ритмом, активируясь около пика каждого цикла, тогда как гиппокампальные и септальные клетки демонстрировали более слабые фазовые предпочтения. Этот сдвиг говорит о том, что во время сновидений глубокие центры бодрствования берут на себя более ритмическую, координирующую роль.
Что это значит для сна, памяти и баланса мозга
В совокупности результаты демонстрируют гибкое, зависящее от состояния сотрудничество между гиппокампом и его подкорковыми партнёрами. В тихих состояниях острые рипплы позволяют гиппокампу посылать мощные, но ограниченные по распространению всплески наружу, вероятно поддерживая реплей памяти, не вызывая при этом значительных изменений в бодрствовании или поведении. В более активных, тета-доминирующих состояниях краткие супрамаммиллярные всплески могут выборочно сдерживать или синхронизировать выход гиппокампа, особенно в CA1, тогда как REM-сон позволяет глубоким нейронам ритмически согласовываться с системами памяти. Для непрофессионального читателя ключевая идея такова: структуры памяти и центры бодрствования постоянно ведут двунаправленные переговоры, подстраивая, кто ведёт, а кто следует при переходе от бодрствования к разным стадиям сна, что помогает сохранять воспоминания и одновременно поддерживать общий баланс состояния мозга.
Цитирование: Caneo, M., Espinosa, N., Lazcano, G. et al. Hippocampo–supramammillary coupling across sleep and wake. Sci Rep 16, 10465 (2026). https://doi.org/10.1038/s41598-026-37066-0
Ключевые слова: гиппокамп, сон и память, ритмы мозга, супрамаммиллярное ядро, нейронная коммуникация