Каждый день мы без усилий замечаем закономерности: красный свет означает «стоп», многолюдная улица — ехать медленнее, определённая поза животного — что оно вот-вот прыгнёт. За этими навыками стоит способность мозга выявлять скрытую, или «латентную», структуру мира и повторно использовать её в самых разных задачах. В этой статье задаётся на первый взгляд простой вопрос: почему один паттерн популяционной активности мозга лучше другого для быстрого и точного решения многих связанных задач?
Скрытые регуляторы, стоящие за нейронными кодами Figure 1.
Авторы исследуют активность мозга на уровне популяций, рассматривая спайки многих нейронов как точки в высокоразмерном пространстве. В фокусе — задачи, которые разделяют набор общих латентных переменных — например форма, размер и положение объекта или положение и скорость животного. downstream-нейрон или схема считывает эти паттерны простым линейным правилом, подобно тому как провести плоскость через облако точек, чтобы отделить «категорию A» от «категории B». Вместо симуляции каждого нейрона в деталях, авторы выводят аналитическую формулу, предсказывающую, насколько хорошо такой считыватель будет обобщать на новые примеры, исходя из геометрии нейронной активности. Удивительно, но выясняется, что поведение определяется всего четырьмя статистиками, которые отражают, насколько сильно нейроны кодируют латентные переменные, насколько чётко разные переменные разнесены, как устроен шум и сколько эффективных размерностей занимает активность.
Четыре простых ингредиента хорошего обобщения
Первый ингредиент — общая корреляция между отдельными нейронами и латентными переменными: когда малые изменения скрытых переменных вызывают заметные сдвиги в нейронных ответах, у downstream-считывателей больше сигнала для работы. Второй и третий ингредиенты описывают «факторизацию»: в идеале разные латентные переменные кодируются вдоль независимых направлений, а случайный шум меняется в направлениях, ортогональных этим сигнальным осям. Это облегчает перенос единой линейной границы через множество задач, которые все зависят от одной и той же скрытой структуры. Четвёртый ингредиент — эффективная размерность, которая отражает, сколько направлений в пространстве активности популяция действительно использует. Более высокая размерность имеет тенденцию растягивать шум по большему числу направлений, повышая надёжность, но это должно быть сбалансировано с тем, насколько чётко сигнал выровнен по поведенчески релевантным переменным.
Проверка теории в искусственных и биологических мозгах Figure 2.
Чтобы проверить свою теорию, авторы сначала применяют её к искусственным нейронным сетям. В многослойных перцептронах, обученных на множестве связанных задач классификации, и в глубокой сети, обученной отслеживать части тела мыши на видео, они измеряют четыре геометрические величины на каждом слое. Предсказанные ошибки тесно совпадают с фактической производительностью простых считывателей, обученных на этих внутренних представлениях. Затем они переходят к данным реального мозга. Записи из зрительных областей макаки показывают, что по мере продвижения сигналов от глаз через более высокие уровни зрительной коры геометрия изменяется таким образом, что снижает ошибку обобщения: корреляции с латентными переменными растут, бессущевая вариабельность отталкивается от сигнальных направлений, а определённые формы размерности перераспределяются. У крыс, обучающихся задаче пространственной альтерации, и поведение, и эффективность считывателя улучшаются за дни тренировки, в то время как геометрия активности гиппокампа и префронтальной коры меняется систематично, в духе предсказаний теории.
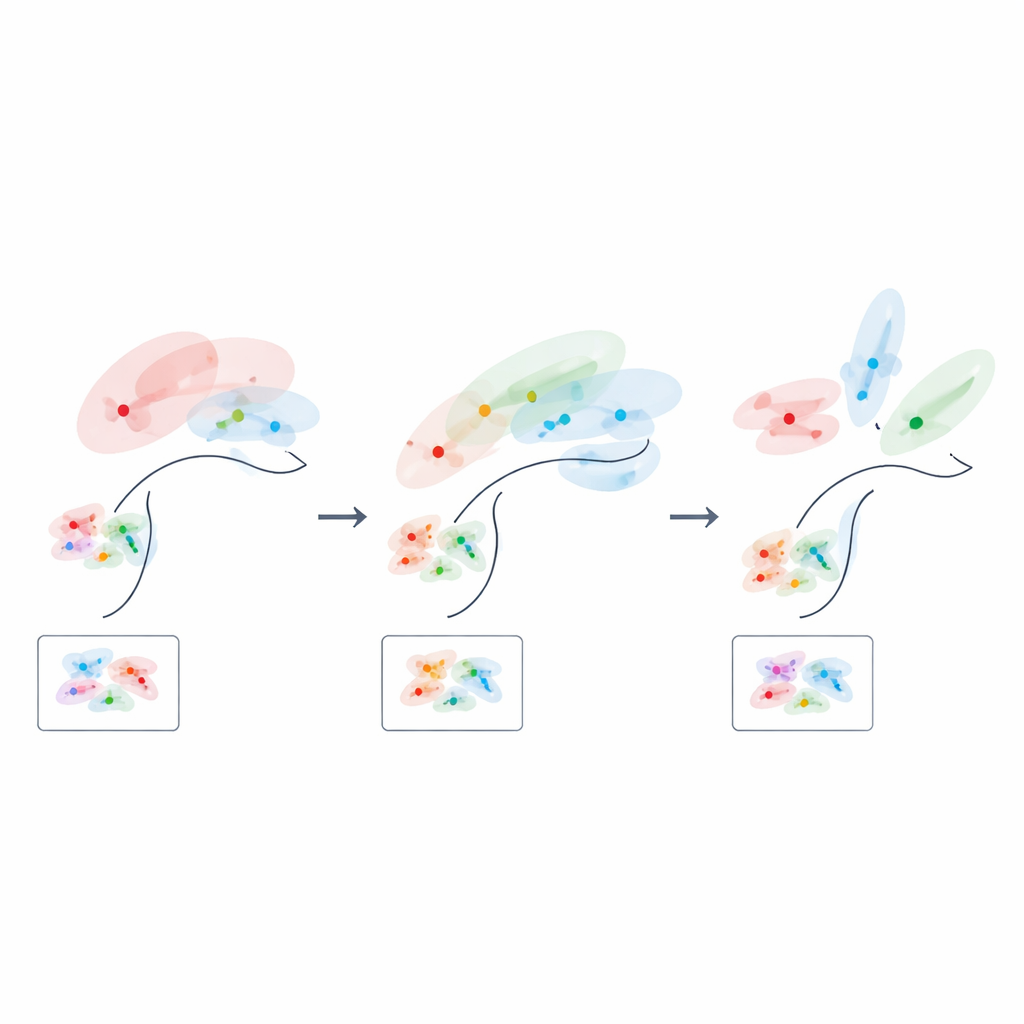
Как обучение переписывает нейронное пространство
Поскольку их формула напрямую связывает геометрию с производительностью, авторы могут спросить, каким должно быть «оптимальное» нейронное кодирование на разных этапах обучения. На раннем этапе, когда доступно лишь несколько обучающих примеров, лучшие коды низкоразмерны и сильно выровнены с наиболее информативными латентными переменными, эффективно сжимая менее полезные признаки. По мере накопления опыта оптимальное решение смещается: представление задачово-важной структуры расширяется в большее число размерностей, и тесная корреляция между отдельными нейронами и отдельными переменными фактически ослабляется. Иными словами, мозг, по-видимому, начинает с сфокусированного, низкоразмерного наброска задачи и постепенно заполняет его более богатой, распределённой картой по мере обучения.
Почему это важно для понимания мозга и машин
Для неспециалиста главное сообщение в том, что популяционная активность мозга — это не просто клубок спайков; у неё есть форма, и эта форма имеет значение. Выделив четыре измеримые геометрические характеристики, которые контролируют, насколько хорошо простые считыватели могут обобщать между связанными задачами, эта работа предлагает общий язык для сравнения биологических и искусственных нейронных сетей. Она указывает на то, что по мере обучения животные и машины реорганизуют свою внутреннюю активность от компактных, сильно выровненных кодов в сторону более высокоразмерных, лучше факторизованных представлений, которые при этом защищают релевантную для задачи информацию от шума. Этот геометрический взгляд помогает объяснить, как одни и те же мозговые цепи гибко повторно используют скрытую структуру в разных ситуациях, обеспечивая то самое кажущееся лёгким обобщение, лежащее в основе повседневного интеллекта.
Цитирование: Wakhloo, A.J., Slatton, W. & Chung, S. Neural population geometry and optimal coding of tasks with shared latent structure.
Nat Neurosci29, 682–692 (2026). https://doi.org/10.1038/s41593-025-02183-y
Ключевые слова: геометрия нейронной популяции, кодирование латентных переменных, обучение многозадачности, развёрнутые представления, обобщение в нейронных сетях