Clear Sky Science · ru
Сорорин запирает выходные ворота ДНК в коисине, чтобы сохранять сцепление сестринских хроматид
Держать скопированные хромосомы вместе в безопасности
Каждый раз при делении клетки она должна скопировать свою ДНК и затем передать по одному полному набору каждой дочерней клетке. Если эти копии разойдутся слишком рано, это может привести к разрывам хромосом, генетическому хаосу и заболеваниям, включая рак. В этом исследовании показано, как небольшая часть белка — хвост Сорорина — действует как физический замок на кольцевой скобе, которая удерживает сестринские копии ДНК вместе, объясняя, как клетки сохраняют пары хромосом до точного момента разделения.
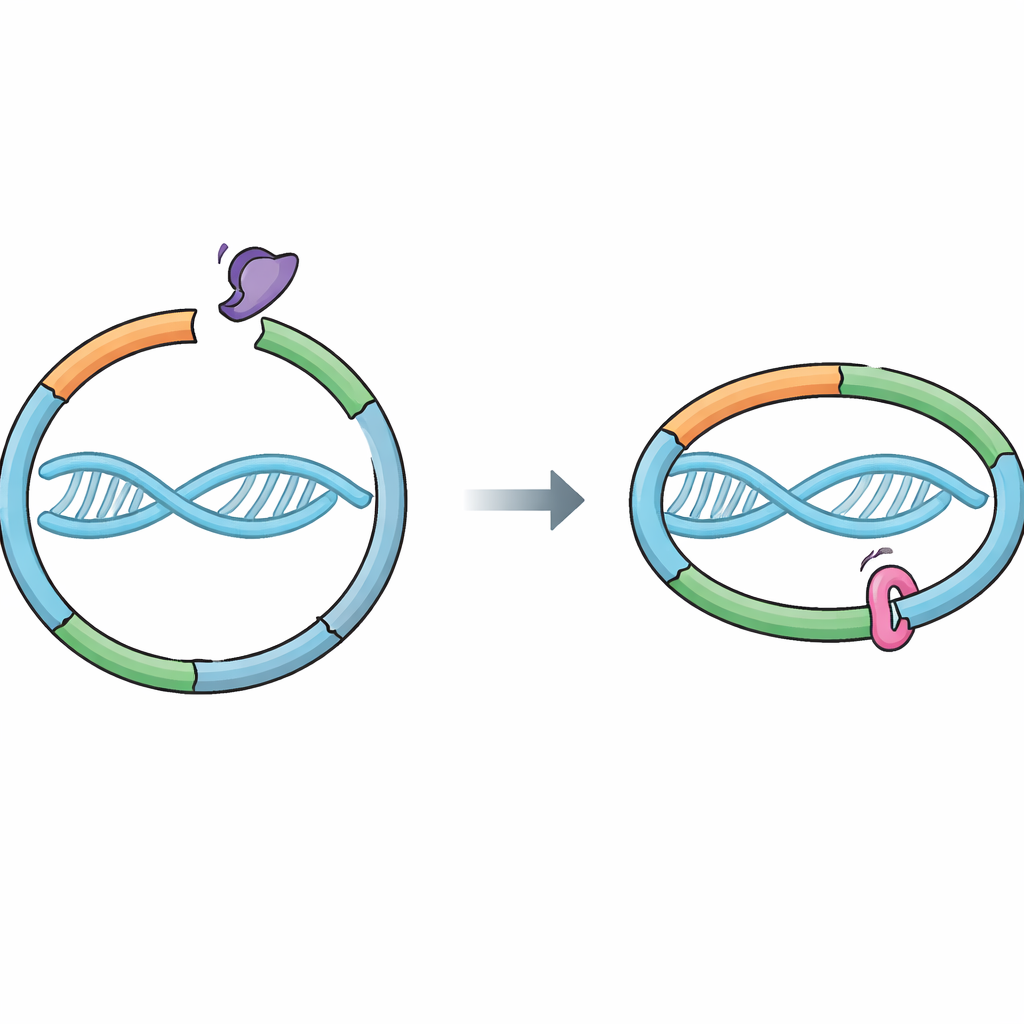
Молекулярное кольцо, охраняющее наши гены
Клетки полагаются на белковый аппарат под названием кокезин, который удерживает вновь скопированные хромосомы вместе. Кокезин формирует кольцо, обхватывающее ДНК и связывающее две сестринские нити, образованные во время репликации. Эта связь должна быть достаточно прочной, чтобы выдержать все встряски деления клетки, но при этом размыкаться в нужный момент, чтобы сестры могли разойтись без повреждений. Белок-помощник Вапл способствует открытию специфических «выходных ворот» в кольце, где встречаются две части кокезина — RAD21 и SMC3. Открытие этих ворот позволяет ДНК выскользнуть, и кокезин слетает. До настоящего времени считалось, что Сорорин сохраняет сцепление главным образом блокируя партнёра Вапла — Pds5, однако оставалось неясным, касается ли Сорорин непосредственно кольца кокезина.
Крошечный хвост с большой держательной силой
Авторы сосредоточились на крайнем хвостовом фрагменте Сорорина — последних 30 аминокислот, называемых С-терминальной областью (CTR). Укорачивая Сорорин и испытывая его фрагменты в человеческих клетках, они обнаружили, что этот крошечный CTR одновременно необходим и достаточен для сохранения сцепления сестринских хромосом. Когда CTR искусственно закрепляли в центромерах — центральных участках, где сестры держатся дольше всего — он практически полностью восстанавливал сцепление в клетках, лишённых полного Сорорина. Напротив, при закреплении CTR по всей хроматине кокезин уже нельзя было эффективно удалить, и хромосомы не могли правильно сжаться и распутаться. Такие клетки демонстрировали длинные, пушистые митотические хромосомы, ДНК‑мосты между расслаивающимися сестрами и ошибки сегрегации — явные симптомы, очень похожие на те, что наблюдаются при удалении самого Вапла.
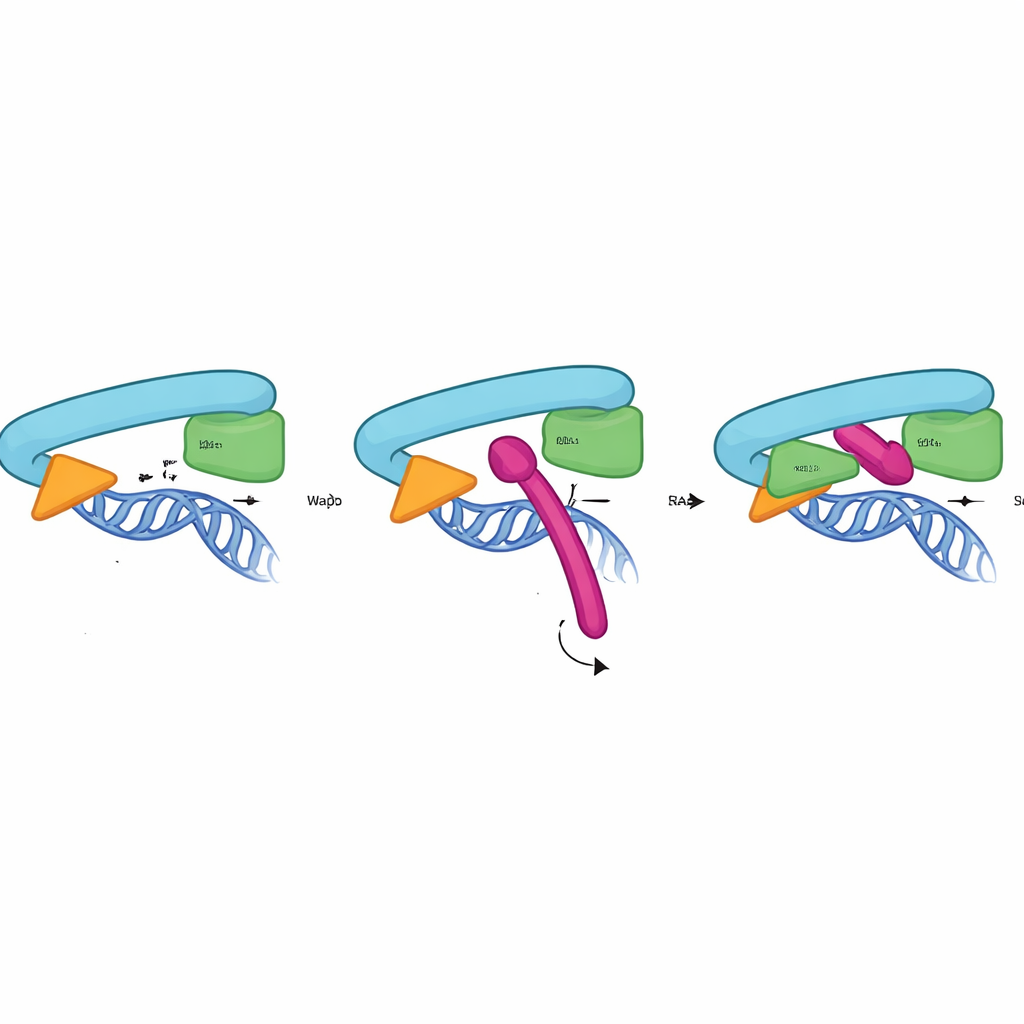
Как замок Сорорина захватывает кольцо
Чтобы понять работу CTR на атомном уровне, исследователи объединили биохимические тесты, мутагенез и предсказания структуры с помощью AlphaFold3. Они показали, что CTR связывается прямо и специфично с интерфейсом между RAD21 и головным доменом SMC3 — тем самым местом, которое формирует выходные ворота для ДНК. Это взаимодействие не требует Pds5 или субъединицы SA2 и возникает только на связанном с хроматином кокезине, достигая максимума в фазе, когда сцепление должно быть стабильно поддержано. Структурная модель указывает, что короткая спираль внутри CTR вклинивается в карман, образованный RAD21 и SMC3, используя набор консервативных гидрофобных и отрицательно заряженных остатков для сцепления. Мутация лишь нескольких точек контакта в Сорорине, RAD21 или SMC3 была достаточна, чтобы устранить связывание и вызвать преждевременное расхождение сестринских хроматид, что подтверждает: этот интерфейс действует как физический замок ворот.
Временной переключатель для разделения хромосом
Клеткам также нужно вовремя освобождать кокезин. Команда обнаружила, что у Сорорина есть соседняя контрольная точка — одна аминокислота (S145), которая фосфорилируется митотической киназой Aurora B. Эта модификация специально нарушает захват Сорорина за Pds5, позволяя Ваплу вновь включиться и способствовать удалению кокезина с плеч хромосом, при этом контакт CTR с воротами RAD21–SMC3 остаётся в значительной степени нетронутым. В центромерах другой фактор, Sgo1, привлекает фосфатазу, снимающую этот фосфат, что сохраняет связь Сорорин–Pds5 и прочное запирание ворот до окончательного разреза RAD21 в анафазе. Генетические тесты показали, что при удалении Вапла дефекты сцепления, вызванные нарушением взаимодействия CTR с воротами, исчезают, подчёркивая, что ключевая роль Сорорина — защищать выходные ворота именно от открывания, вызываемого Ваплом.
Что это значит для стабильности генома и болезней
Вместе эти результаты перестраивают представление о Сорорине как о защитнике с двойной функцией: он одновременно конкурирует с Ваплом за доступ к Pds5 и прямо зажимает выходные ворота кокезина для ДНК через свой крошечный CTR. Эта двойная регуляция обеспечивает прочное сцепление там, где это нужно, особенно в центромерах, и одновременно даёт возможность снять сцепление с плеч хромосом в начале митоза, чтобы обеспечить правильное сжатие и распутывание. Поскольку мутации в кокезине и его регуляторах часто встречаются при нарушениях развития и раке, точно нанесённая контактная поверхность между Сорорином, RAD21 и SMC3 выделяет новую уязвимость, где небольшие изменения могут дестабилизировать хромосомы — и потенциальную мишень для будущих терапий, стремящихся тонко регулировать степень удержания нашего генетического материала.
Цитирование: Chen, Q., Yuan, X., Shi, M. et al. Sororin locks the DNA-exit gate of cohesin to preserve sister-chromatid cohesion. Nat Commun 17, 2284 (2026). https://doi.org/10.1038/s41467-026-70484-2
Ключевые слова: сцепление сестринских хроматид, комплекс кокезин, Сорорин, сегрегация хромосом, стабильность генома