Clear Sky Science · ru
Мультивалентный адаптерный механизм обеспечивает импорт протеасом в ядро
Как клетки поддерживают чистоту в своих «пультовых»
Внутри каждой клетки ядро выполняет роль командного центра, заполненного ДНК и белками, контролирующими включение и выключение генов. Чтобы этот центр работал исправно, клетки полагаются на мощные молекулярные «шредеры» — протеасомы, разрушающие изношенные или неисправные белки. В этой работе показано, как клетки решают, на первый взгляд непростую задачу — транспортировать массивные протеасомы через узкие ворота в ядро в нужный момент.
Проблема перемещения больших «машин»
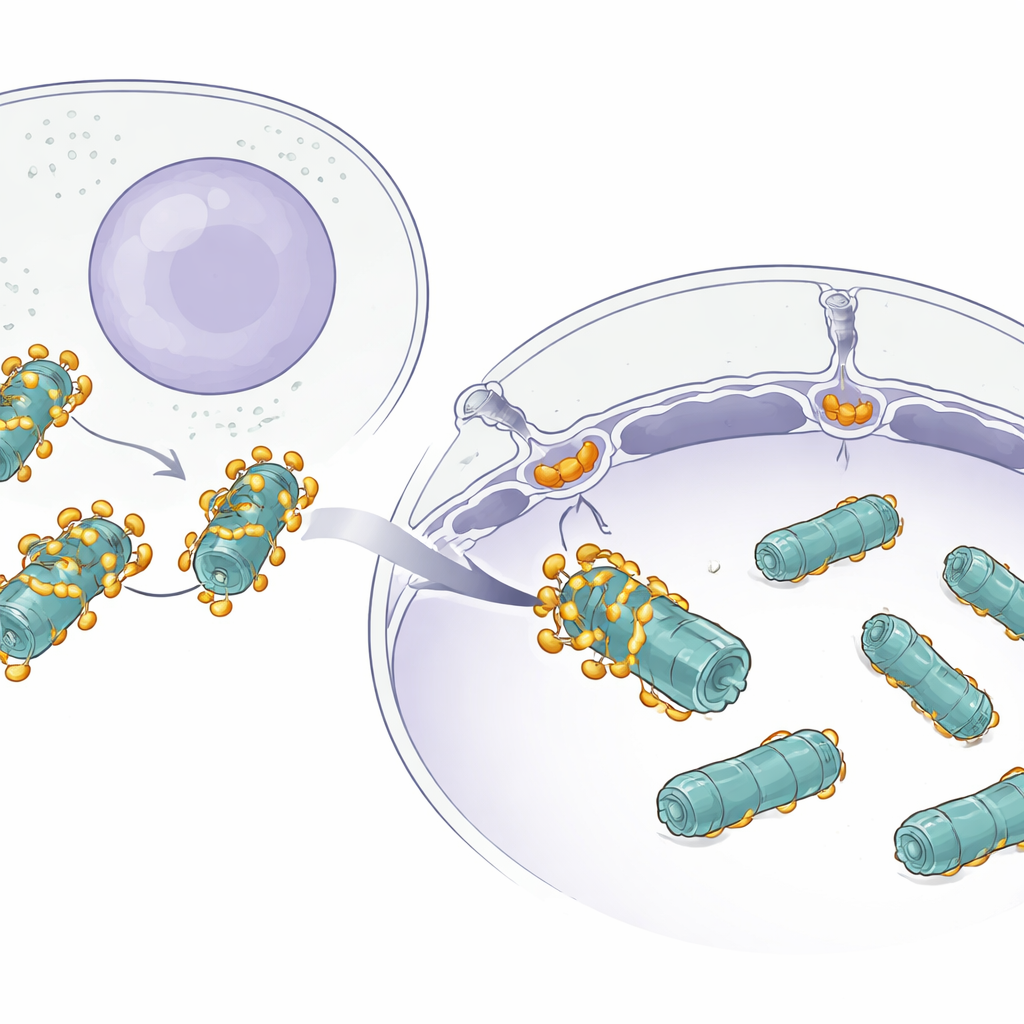
Многие белки тихо перемещаются между цитоплазмой и ядром через гигантские порты в ядерной оболочке, называемые ядерными порами. Маленькие белки проходят легко, но большие комплексы, такие как протеасомы, — другое дело: это массивные, бочкообразные структуры, собранные из множества частей. Ученым было известно, что небольшие грузы используют помощников — импортинов — для проникновения в ядро, но было непонятно, как нечто столь крупное, как протеасома, проходит эффективно, не застревая и не теряясь.
Помощник, меняющий форму, выходит на сцену
Группа сосредоточилась на небольшом гибком белке AKIRIN2, который предыдущие исследования показали как абсолютно необходимый для проникновения протеасом в ядро. Клетки, делящиеся без AKIRIN2, не могут доставлять протеасомы в ядро и быстро гибнут из‑за накопления поврежденных ядерных белков. Сочетая разные методы — масштабное сканирование мутаций, затрагивающее почти каждую аминокислоту AKIRIN2, высокоразрешающую криоэлектронную микроскопию для 3D‑визуализации комплекса и тщательные биохимические испытания — исследователи сопоставили, какие участки AKIRIN2 важны и каковы их функции. Они обнаружили ранее недооцененный «крыловидный» сегмент и спиральный участок, который позволяет AKIRIN2 образовывать димеры (пары), — оба играют ключевые роли в захвате протеасом и других партнёров.
Создание многохватальной транспортной рукояти
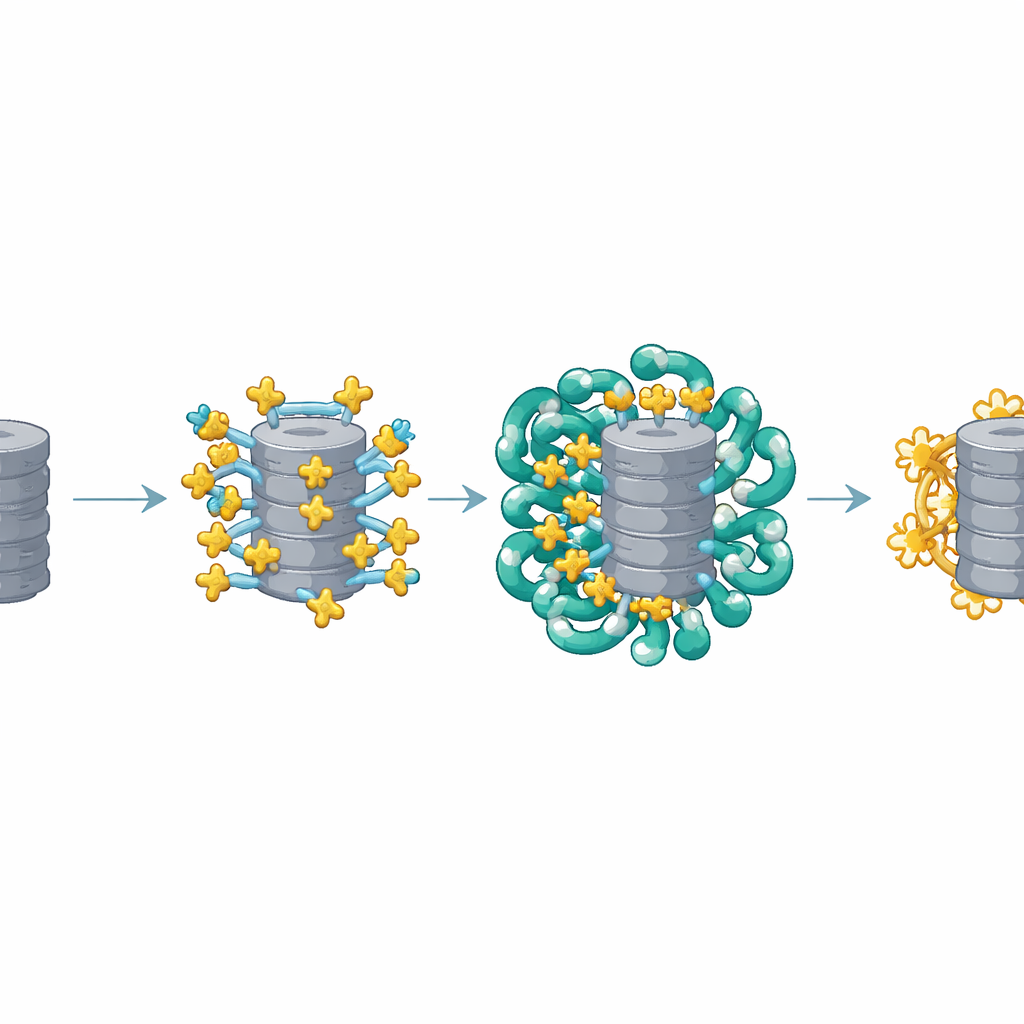
Структурные изображения показали, что несколько копий AKIRIN2 собираются на одной стороне протеасомы, словно множество рук хватают один объект. Каждая пара AKIRIN2 связывается с определёнными участками на внешней поверхности протеасомы и одновременно взаимодействует с разными импортимами. Один из импортинов, IPO9, удерживает AKIRIN2 необычным образом, обвивая его и меняя конформацию при сборке комплекса. Другие импортиновые пары распознают короткие сигналы ядерной локализации, несомые AKIRIN2. Поскольку на одной протеасоме расположено несколько молекул AKIRIN2, они коллективно демонстрируют множество таких сигналов одновременно. Эта мультивалентная «рукоять» позволяет толпе импортинов прикрепляться синхронно, обеспечивая достаточную силу, чтобы протеасома прошла через ядерную пору.
Разумный цикл сборки и освобождения
Реконституционные эксперименты в упрощённой пробирочной системе показали, что протеасомы эффективно попадают в ядро только при одновременном наличии AKIRIN2 и импортинов. Чем больше AKIRIN2, тем сильнее и быстрее импорт. Оказавшись в ядре, транспортный аппарат должен быть удалён, чтобы протеасомы могли выполнять свою работу. Для этого клетка использует двухступенчатое освобождение. Небольшой переключающий белок Ran в форме, связанной с GTP, снимает большинство импортинов со груза. AKIRIN2 и часть IPO9 держатся более упрямо, но у ядра есть запасной механизм: ядерные протеасомы непосредственно разрушают сам AKIRIN2, без обычной метки убиквитином. Измерения в разные фазы клеточного цикла показывают, что уровень AKIRIN2 достигает максимума во время деления, когда формируются новые ядра, а затем падает в ранней G1, что согласуется с этим целенаправленным удалением.
Почему это важно не только для одного белка
В совокупности эти результаты показывают, что AKIRIN2 действует как гибкий каркас, собирающий множество импортинов на одной протеасоме и превращающий сложную задачу транспорта в скоординированное групповое усилие. Кластеризуя сигналы для входа в ядро на адаптере, а не на самой протеасоме, клетки могут контролировать, когда и с какой интенсивностью крупные комплексы перемещаются в ядро, не перестраивая эти комплексы. Похожие адаптерные стратегии, по-видимому, используются и для других крупных белковых сборок, а некоторые вирусы эксплуатируют родственные приёмы для вторжения в ядро. Понимание этой общей логики транспорта углубляет наше представление о том, как клетки поддерживают баланс белков в самой чувствительной компартменте, и в перспективе может указать пути вмешательства при нарушениях разложения белков или функции ядра.
Цитирование: Brunner, H.L., Kalis, R.W., Grundmann, L. et al. A multivalent adaptor mechanism drives the nuclear import of proteasomes. Nat Commun 17, 2359 (2026). https://doi.org/10.1038/s41467-026-69162-0
Ключевые слова: ядерный транспорт, протеасома, AKIRIN2, импортин, разрушение белков