Clear Sky Science · ru
Динамический отпечаток компромисса активность–стабильность в эволюции лактамаз
Почему это важно для антибиотикорезистентности
Антибиотикорезистентность часто кажется чёрным ящиком: бактерии «как‑то» изменяются, и лекарства перестают работать. Это исследование приоткрывает этот ящик для одного из наиболее известных ферментов устойчивости — TEM‑1 β‑лактамазы, которая помогает бактериям разрушать антибиотики типа пенициллина. Наблюдая, как форма и движения этого фермента меняются по мере его эволюции в ответ на новые препараты, авторы показывают, почему мутации, повышающие активность, зачастую ухудшают стабильность, и как эволюция находит хитрые пути для балансировки обоих свойств.
От борца с пенициллином до разрушителя более широкого спектра препаратов
Изначально TEM‑1 превосходно разлагает старые пенициллиновые антибиотики, но ему трудно справляться с более объёмными современными препаратами, такими как цефотаксим. Во многих клинических штаммах рядом с активным центром — карманом, где антибиотики расщепляются — встречается одна ключевая мутация G238S. Эта мутация резко улучшает способность фермента разрушать цефотаксим, при этом лишь умеренно ухудшая его исходную активность по отношению к пенициллину. Авторы показывают, что G238S не просто расширяет карман; она перестраивает то, как движутся несколько окружающих петель и спиралей, создавая новую функциональную конформацию, которая лучше вмещает объёмные молекулы.
Движения белка как регулятор эволюционной настройки
Используя продвинутые методы ЯМР (ядерного магнитного резонанса), исследователи измеряли, как разные участки TEM‑1 двигаются на временнЫх шкалах от триллионных до тысячных долей секунды. Дикий тип TEM‑1 довольно жёсткий, что помогает ему эффективно обрабатывать свои первоначальные субстраты. G238S сохраняет большую часть этой быстрой жёсткости, но вводит медленные, тщательно настроенные движения во многих стенках активного сайта. Эти движения достаточно быстры, чтобы укладываться в темп химии фермента, но не настолько сильны, чтобы нарушать критические каталитические остатки. В результате возникает «оптимальное окно» гибкости: чуть‑чуть движение, чтобы открыть карман для цефотаксима, при этом сохраняя выровненность основной химической машины.
Баланс двух форм вместо выбора одной
Эволюция не замораживает TEM‑1 в одной новой форме. Вместо этого фермент образует по крайней мере две основные конформации: состояние «пенициллиназы», похожее на исходную структуру, и более открытое состояние «цефотаксимазы», лучшее для новых препаратов. Последующие мутации, такие как E104K и A42G, действуют тонко. Вместо того чтобы просто стабилизировать новую, благоприятную для цефотаксима форму, они перестраивают соотношение между двумя состояниями. Данные ЯМР показывают, что разные части активного центра и опорного каркаса могут сдвигать свои популяции независимо вдоль этого двухсостояния континуума. Это создаёт комбинаторное множество вариантов фермента, каждый с разной смесью пенициллиноподобных и цефотаксимоподобных конформаций и, следовательно, с разными каталитическими профилями.
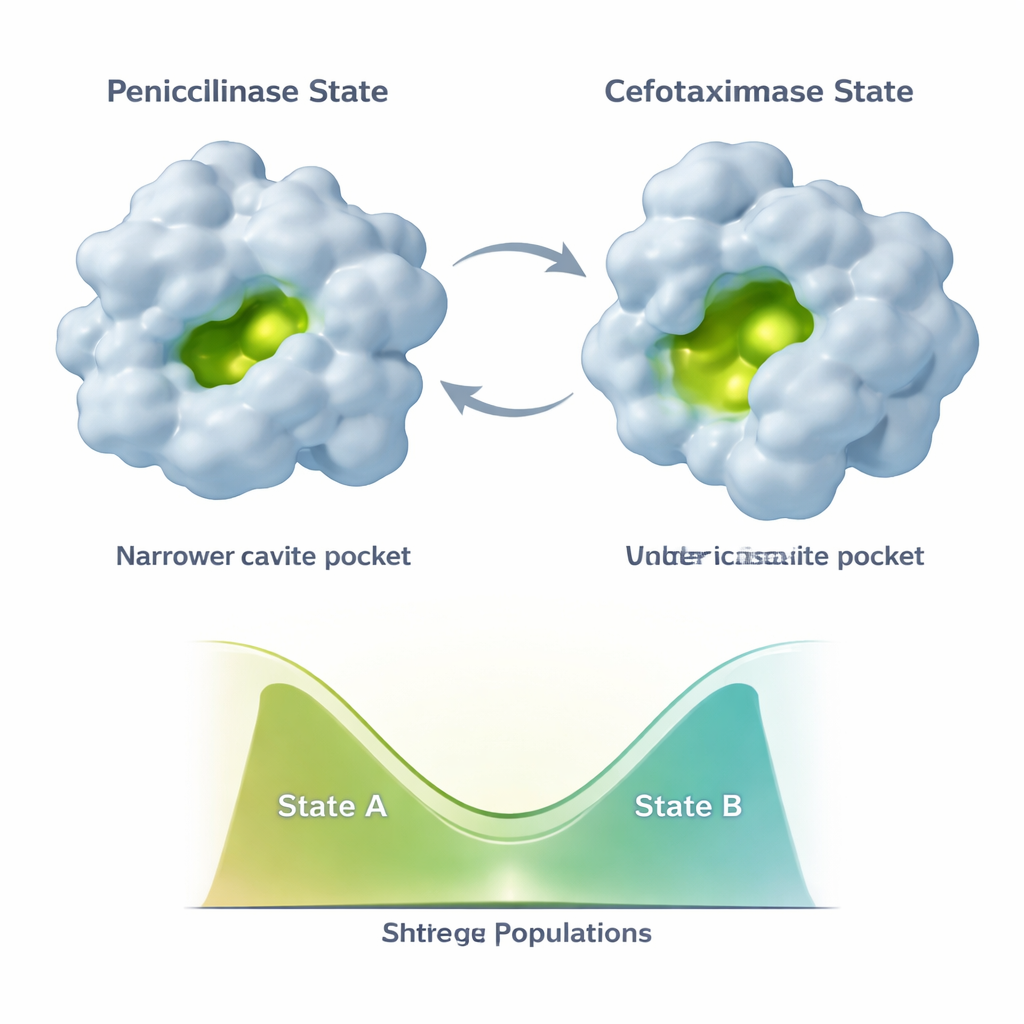
Скрытые слабые места и отдалённые «починки»
Мутации, улучшающие активность, часто несут скрытую цену: они делают белок менее стабильным. Вместо того чтобы смотреть только на полное разворачивание, команда картировала локальную стабильность на уровне коротких сегментов с помощью гидрид-дейтериевого обмена и масс‑спектрометрии. Выяснилось, что G238S дестабилизирует не только близлежащие петли, но и отдалённые спирали и β‑листы, формирующие структурный «скелет». Некоторые из этих областей перекрываются с «криптическим» аллостерическим карманом — редко посещаемым отверстием в ядре белка, которое может связывать малые молекулы и подавлять активность. G238S упрощает открывание этого кармана, фактически встраивая в фермент слабую самоингибирующую черту. Последующие мутации, особенно A42G, укрепляют этот ослабленный каркас, повышая локальную стабильность вокруг трёх взаимодействующих спиралей, не устраняя при этом полезную динамику активного сайта. Иными словами, эволюция латает отдалённые структурные слабые места, не отменяя первоначальную инновацию.
Что исследование говорит о стратегии эволюции
Для непрофессионального читателя главный вывод в том, что белки вроде TEM‑1 не приобретают резистентность простым переключением «вкл/выкл». Каждая мутация слегка перекраивает то, как фермент «дышит», гибко меняет форму и распределяет время между разными рабочими позами. G238S открывает дорогу новой функции — разрушению современных антибиотиков — но также создаёт локальные уязвимости и частично самоингибирующее состояние. Секундные мутации работают как осторожные усиления, стабилизируя каркас и тонко настраивая баланс между старыми и новыми конформациями, чтобы фермент оставался одновременно активным и прочным. Этот динамический взгляд на эволюцию, где движения и локальные слабости важны не меньше, чем статическая структура, может помочь в разработке будущих антибиотиков и препаратов, нацеленных на ферменты, которые будет труднее обойти бактериям.
Цитирование: Arcia, E., Keramisanou, D., Jacobs, L.M.C. et al. Dynamic signature of activity-stability tradeoff in lactamase evolution. Nat Commun 17, 1884 (2026). https://doi.org/10.1038/s41467-026-68620-z
Ключевые слова: антибиотикорезистентность, бета-лактамаза, эволюция белков, динамика ферментов, стабильность белков