Clear Sky Science · ru
Молекулярная основа антагонизма димерного человеческого рецептора аргининового васопрессина V1A
Почему важен гормональный рецептор в мозге
Гормоны, такие как васопрессин и окситоцин, наиболее известны контролем водного баланса, кровяного давления, родовой деятельности и привязанности. Тем не менее механизмы работы их рецепторов на атомном уровне оставались во многом скрытыми. В этой работе представлены детальные трёхмерные структуры одного ключевого рецептора — человеческого васопрессинового рецептора V1a, связанного с социальным поведением, стрессом и рядом заболеваний мозга. Понимание его формы и того, как препараты блокируют этот рецептор, может помочь учёным разработать лучшие методы лечения таких состояний, как аутизм, посттравматическое стрессовое расстройство и болезнь Хантингтона.
Двойной рецептор, формирующий сигналы для сердца, почек и мозга
Рецептор V1a находится на поверхности многих клеток организма, особенно в кровеносных сосудах, почках и отдельных участках мозга. Когда на него садится гормон васопрессин, рецептор запускает внутренние сигнальные пути, контролирующие кровяное давление, водный баланс и мозговые цепи, отвечающие за социальное взаимодействие, эмоции и стресс. Генетические и клинические исследования связывают ненормальную передачу сигналов через V1a с расстройствами аутистического спектра, ПТСР и болезнью Хантингтона, что делает его привлекательной мишенью для лекарств. Несколько блокаторов V1a (антагонистов) уже используются или проходят клинические испытания, но до сих пор никто не видел человеческий V1a с высоким разрешением, что оставляло открытыми ключевые вопросы о том, как он собирается и как именно эти препараты его выключают. 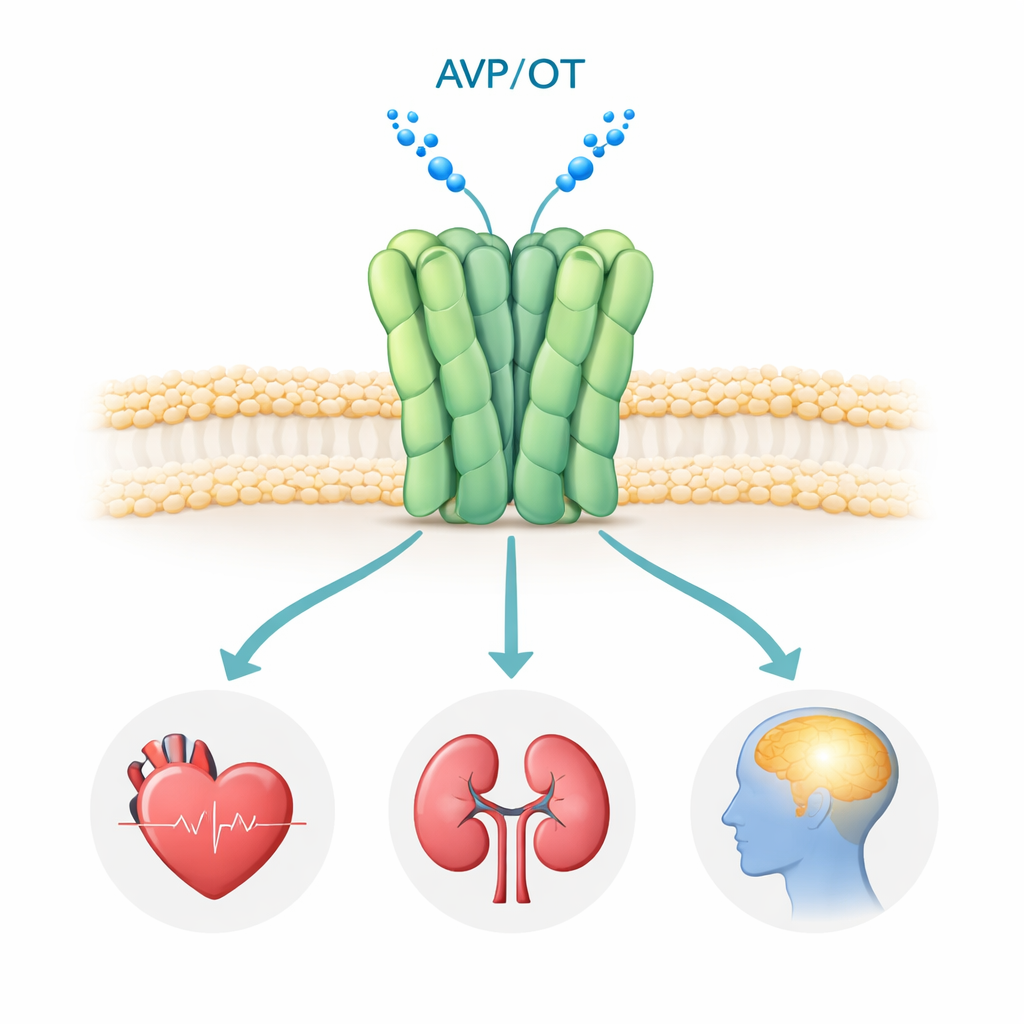
Фиксация структуры рецептора в нескольких состояниях, связанных с препаратами
Исследователи использовали крио-электронную микроскопию — метод, который быстро замораживает белки и снимает их с помощью электронного луча — чтобы визуализировать человеческий рецептор V1a. Чтобы стабилизировать белок, они создали слегка модифицированную форму, которая по-прежнему хорошо связывает препараты, и присоединили к ней небольшой фрагмент антитела (нанотело) для помощи в визуализации. Были получены структуры рецептора в апо-состоянии и в комплексе с тремя медицински важными антагонистами: атосибаном (пептидный препарат, используемый для предотвращения преждевременных родов) и двумя проникающими в мозг малыми молекулами, баловавптаном и SRX246, которые тестировались у людей с аутизмом или болезнью Хантингтона. Все структуры достигли почти атомного разрешения, раскрывая расположение семи спиралей, проходящих через мембрану, гибких петель и связанных препаратов.
Рецептор, который предпочитает работать парами
В отличие от родственных рецепторов, которые ранее наблюдали только как одиночные молекулы, V1a во всех четырёх структурах крио-ЕМ оказался парой — димером. Два рецептора лежат рядом в мембране, устанавливая плотные контакты главным образом через одну из своих спиралей, с участием как полярных, так и гидрофобных взаимодействий. Чтобы проверить, встречается ли такое парование в живых клетках, команда слила V1a с ярким флуоресцентным белком и использовала метод фотоблекания одиночных молекул: если в точке на поверхности клетки было два копий рецептора, свет исчезал в два шага. Примерно три четверти наблюдаемых пятен обесцвечивались ровно в два шага, что убедительно подтверждает идею о том, что V1a естественно образует димеры на поверхности клетки. Когда учёные мутировали ключевые остатки, ответственные за контакты, нарушая интерфейс, рецептор смещался в сторону одиночных единиц и становился менее чувствительным к гормону и препарату, что указывает на то, что димер не является просто структурной деталью, а функционально важен. 
Гибкие ворота у входа гормона
Команда обнаружила неожиданную «воротную» область, названную внеклеточной петлёй 2 (ECL2), которая расположена в верхней части кармана связывания гормона. В состоянии без препарата (апо) эта петля лежит плоско над карманом, подобно крышке, и не образует обычную дисульфидную связь (связь серы с серой), которую наблюдают во многих родственных рецепторах. Вместо этого части петли загибаются внутрь кармана и удерживаются сетью взаимодействий с окружающими спиралями, частично закрывая большой «липкий» карман связывания. Когда связывается любой из трёх антагонистов, ECL2 поднимается и отводится в сторону, формирует классическую дисульфидную связь и создаёт широкую полость, заполненную растворителем, которую заполняют препараты. Это драматическое движение указывает на то, что V1a может использовать ECL2 как динамический барьер, ограничивающий случайную активацию посторонними молекулами, и что препараты можно проектировать либо чтобы зафиксировать петлю в её плоском «основном» состоянии, либо чтобы воспользоваться её поднятой, открытой конформацией.
Как три препарата заглушают один и тот же рецептор разными путями
Атосибан, который тесно имитирует природный гормон окситоцин, простирается от вершины кармана до его основания, закрепляясь за счёт комбинации водородных связей и гидрофобных контактов. Изменяя несколько ключевых позиций по сравнению с окситоцином, он не вызывает цепочки внутренних сдвигов, обычно необходимых для активации рецептора: критические «микровыключатели», которые перемещаются во время передачи сигнала, остаются зафиксированными в неактивных положениях, внутренняя полость, допускающая G-белок, не раскрывается, а сайт связывания магния, важный для активации, нарушается. Напротив, баловавптан и SRX246 — компактные непептидные молекулы, которые проникают глубоко в карман, но используют разные стратегии. Баловавптан опирается на жёсткое гидрофобное ядро, плотно упаковывающееся в глубокую щель, и гибкий полярный хвост, достигающий входа в карман. SRX246 использует модульную, фрагментоподобную архитектуру, заякоренную β-лактамным сердечником, с разными «зонами», заполняющими субкарманы и простирающимися к внеклеточным петлям. В обоих случаях препараты стабилизируют неактивную конформацию, несовместимую со связыванием G‑белка. Тонкие различия в форме и химии кармана — особенно в двух позициях на спиралях 5 и 7 — помогают объяснить, почему баловавптан и SRX246 предпочитают V1a по сравнению с близкородственными рецепторами.
Значение для будущих терапий
Представляя высокоразрешающие снимки V1a в виде димера, раскрывая ранее не виденную «плоскую» конформацию петли в состоянии без препарата и детализируя, как три очень разные антагониста выключают рецептор, эта работа даёт разработчикам лекарств точную структурную карту для нацеливания на V1a. Она предлагает пути создания препаратов следующего поколения, которые либо используют особенности, специфичные для димера, либо фиксируют рецептор в особенно неактивном основном состоянии, с конечной целью лечить расстройства мозга и связанные со стрессом состояния более прицельно и с меньшими побочными эффектами.
Цитирование: Zhong, P., Chu, B., Yu, Z. et al. Molecular basis of antagonism of the dimeric human arginine vasopressin receptor 1A. Nat Commun 17, 1622 (2026). https://doi.org/10.1038/s41467-026-68331-5
Ключевые слова: рецептор васопрессина V1a, сопряжённый с G-белком рецептор, димеризация рецептора, структура методом крио-ЭМ, дизайн нейропсихиатрических препаратов