Clear Sky Science · pt
Base molecular do antagonismo do receptor humano dimérico da vasopressina arginina 1A
Por que um receptor cerebral de hormônio importa
Hormônios como vasopressina e ocitocina são mais conhecidos por controlar o balanço hídrico, a pressão arterial, o parto e o vínculo social. Mas o modo como seus receptores funcionam ao nível atômico permaneceu em grande parte oculto. Este artigo revela estruturas 3D detalhadas de um receptor chave, o receptor humano de vasopressina V1a, que está associado a comportamento social, estresse e vários transtornos cerebrais. Compreender sua forma e como medicamentos o bloqueiam pode ajudar cientistas a projetar tratamentos melhores para condições como autismo, transtorno de estresse pós-traumático e doença de Huntington.
Um receptor gêmeo que molda sinais do coração, rim e cérebro
O receptor V1a fica na superfície de muitas células do corpo, especialmente em vasos sanguíneos, rins e certas regiões do cérebro. Quando o hormônio vasopressina se liga a ele, o receptor ativa vias de sinalização internas que controlam pressão arterial, equilíbrio de fluidos e circuitos cerebrais relacionados à interação social, emoção e estresse. Estudos genéticos e clínicos associaram sinalização V1a anormal ao transtorno do espectro autista, TEPT e doença de Huntington, tornando-o um alvo farmacológico atraente. Vários medicamentos bloqueadores de V1a (antagonistas) já são usados ou estão em ensaios clínicos, mas até agora ninguém havia observado o receptor humano V1a em alta resolução, deixando grandes questões sobre como ele se monta e exatamente como esses fármacos o inibem. 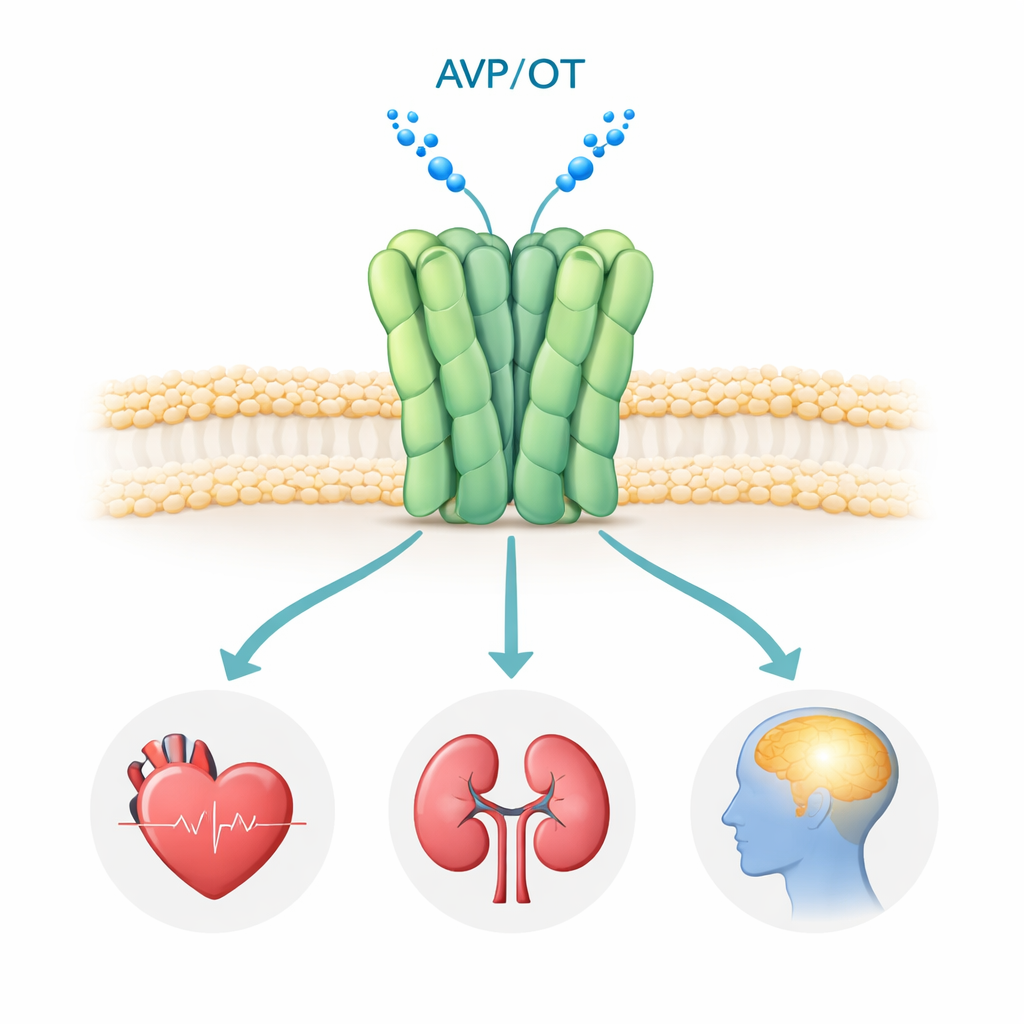
Capturando a estrutura do receptor em múltiplos estados com fármaco ligado
Os pesquisadores usaram crio‑microscopia eletrônica, uma técnica que congela rapidamente proteínas e as imagina com um feixe de elétrons, para visualizar o receptor humano V1a. Para estabilizar a proteína, eles projetaram uma forma ligeiramente modificada que ainda se liga bem a fármacos e a combinaram com um pequeno fragmento de anticorpo (um nanocorpo) para auxiliar a imagem. Resolveram estruturas do receptor isolado e ligado a três antagonistas de relevância médica: atosibano (um peptídeo usado para prevenir parto prematuro) e duas pequenas moléculas capazes de penetrar no cérebro, balovaptan e SRX246, testadas em pessoas com autismo ou doença de Huntington. Todas as estruturas atingiram resolução quase atômica, revelando as posições das sete hélices transmembrana do receptor, laços flexíveis e os fármacos ligados.
Um receptor que prefere atuar em pares
Ao contrário de receptores relacionados que haviam sido vistos apenas como unidades simples, o V1a apareceu como um par — um dímero — em todas as quatro estruturas por crio‑ME. Os dois receptores ficam lado a lado na membrana, fazendo contatos estreitos principalmente por meio de uma de suas hélices, auxiliados por interações tanto polares quanto hidrofóbicas. Para testar se esse emparelhamento também ocorre em células vivas, a equipe fundiu o V1a a uma proteína fluorescente brilhante e usou um método de fotobranqueamento de molécula única: se um ponto na superfície celular continha duas cópias do receptor, sua luz desapareceria em dois passos. Cerca de três quartos dos pontos observados branquearam exatamente em dois passos, apoiando fortemente a ideia de que V1a forma naturalmente dímeros na superfície celular. Quando os cientistas mutaram resíduos de contato chave para perturbar a interface, o receptor mudou para unidades simples e tornou‑se menos responsivo ao hormônio e ao fármaco, sugerindo que o dímero não é apenas decoração estrutural, mas funcionalmente importante. 
Uma porta flexível na entrada do hormônio
A equipe descobriu uma região inesperada de “portão” chamada laço extracelular 2 (ECL2), que fica no topo do bolso de ligação ao hormônio. No estado sem fármaco (apo), esse laço repousa sobre o bolso como uma tampa e não forma a habitual ponte de dissulfeto (uma ligação enxofre‑enxofre) vista em muitos receptores relacionados. Em vez disso, partes do laço se dobram para dentro do bolso e são mantidas no lugar por uma teia de interações com hélices circundantes, cobrindo parcialmente a grande cavidade de ligação pegajosa. Quando qualquer um dos três antagonistas se liga, ECL2 ergue‑se e afasta‑se, forma a clássica ponte de dissulfeto e cria uma ampla cavidade preenchida por solvente que os fármacos ocupam. Esse movimento dramático sugere que V1a pode usar ECL2 como uma barreira dinâmica para limitar ativações aleatórias por moléculas estranhas, e que fármacos poderiam ser projetados tanto para prender o laço em seu estado “plano” quanto para explorar sua conformação elevada e aberta.
Como três fármacos silenciam o mesmo receptor de maneiras diferentes
Atosibano, que imita de perto o hormônio natural ocitocina, estende‑se do topo do bolso até sua base, ancorando‑se por meio de uma combinação de ligações de hidrogênio e contatos hidrofóbicos. Ao mudar algumas posições-chave em comparação com a ocitocina, ele deixa de desencadear a cadeia de deslocamentos internos normalmente exigida para a ativação do receptor: resíduos cruciais “microswitch” que se movem durante a sinalização permanecem travados em suas posições inativas, a cavidade interna que admitiria a proteína G nunca se abre, e um sítio de ligação ao magnésio importante para ativação é perturbado. Em contraste, balovaptan e SRX246 são moléculas pequenas, não peptídicas e compactas que se enterram profundamente no bolso, mas usam estratégias distintas. Balovaptan depende de um núcleo rígido e hidrofóbico que se encaixa firmemente em uma fenda profunda, além de uma cauda polar flexível que alcança a entrada do bolso. SRX246 usa uma arquitetura modular, tipo fragmento, ancorada por um núcleo β‑lactama, com diferentes “zonas” ocupando sub‑bolsos e se estendendo em direção aos laços extracelulares. Em ambos os casos, os fármacos estabilizam uma conformação inativa incompatível com a ligação da proteína G. Diferenças sutis na forma e química do bolso — especialmente em duas posições nas hélices 5 e 7 — ajudam a explicar por que balovaptan e SRX246 preferem V1a em relação a receptores estreitamente relacionados.
Implicações para terapias futuras
Ao fornecer instantâneos em alta resolução de V1a como dímero, revelar uma conformação de laço “plana” não vista antes no estado sem fármaco e detalhar como três antagonistas muito diferentes silenciam o receptor, este trabalho dá aos projetistas de fármacos um mapa estrutural preciso para visar V1a. Ele sugere maneiras de criar medicamentos de próxima geração que exploram características específicas do dímero ou travam o receptor em um estado basal especialmente inativo, com o objetivo final de tratar transtornos cerebrais e relacionados ao estresse de forma mais dirigida e com menos efeitos colaterais.
Citação: Zhong, P., Chu, B., Yu, Z. et al. Molecular basis of antagonism of the dimeric human arginine vasopressin receptor 1A. Nat Commun 17, 1622 (2026). https://doi.org/10.1038/s41467-026-68331-5
Palavras-chave: receptor de vasopressina V1a, receptor acoplado à proteína G, dimerização do receptor, estrutura por crio-ME, projeto de fármacos neuropsiquiátricos