Clear Sky Science · pl
Przejściowa proliferacja przez odwracalną kontrolę YAP i mitogenów nad stosunkiem cykliny D1/p27
Jak tkanki wiedzą, kiedy rosnąć
Gdy przecinasz skórę lub uszkodzisz narząd, pobliskie komórki krótkotrwale przełączają się ze stanu spoczynku w tryb naprawczy, mnożąc się tylko tyle, by załatać ranę, po czym zatrzymują się z powrotem. W artykule postawiono pozornie proste pytanie: jak komórki rozpoznają, kiedy zacząć, a co równie ważne — kiedy przestać się dzielić, żeby gojenie przebiegało bez nadmiernego rozrostu przypominającego nowotwór? Badacze odkrywają wbudowaną grę równowagi wewnątrz komórek, która uruchamia silny sygnał wzrostu tylko tymczasowo, a następnie niezawodnie go wyłącza.
Czerwono‑zielone światło dla podziału komórkowego
W wielu tkankach system molekularny zwany Hippo–YAP działa jak kontroler ruchu dla wzrostu komórek. Gdy tkanki są zatłoczone i nienaruszone, szlak Hippo trzyma białko YAP poza jądrem komórkowym i komórki pozostają w stanie spoczynku. Kiedy tkanka zostaje uszkodzona lub rozciągnięta, Hamowanie Hippo ustaje, YAP przechodzi do jądra i komórki są pobudzane do podziału. Autorzy badali tysiące pojedynczych komórek nabłonkowych hodowanych jako płaskie arkusze, wystawiając je na różne kombinacje czynników wzrostu (mitogenów), leków i różnych gęstości komórek. Stwierdzili, że sygnały od nich pochodzące, fizyczne zagęszczenie i aktywność YAP nie działają w izolacji; zamiast tego zbiegają się na jednym wewnętrznym punkcie decyzyjnym, który determinuje, czy dana komórka ponownie wejdzie w cykl komórkowy.
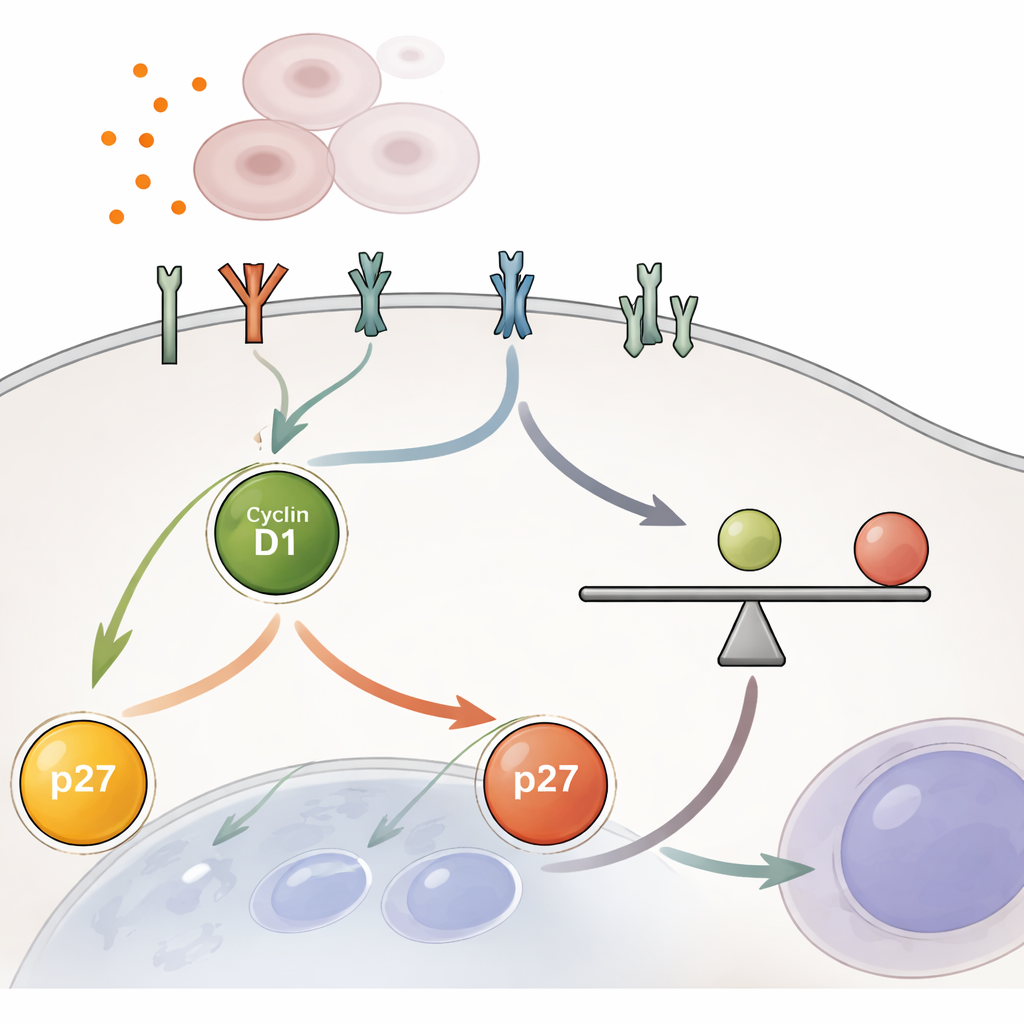
Kluczowy stosunek wewnątrz komórki
Główne odkrycie polega na tym, że decyzja o podziale zapada na podstawie stosunku dwóch białek w jądrze podczas wczesnej fazy wzrostu zwanej G1. Jedno białko, cyklina D1, popycha komórkę ku podziałowi; drugie, p27, działa jak hamulec. Istotny nie jest absolutny poziom któregoś z białek, lecz stosunek cyklina D1/p27. Gdy ten stosunek przekracza krytyczny próg, białko bramkowe Rb zostaje fosforylowane i uwalnia aparat niezbędny do replikacji DNA, pozwalając komórce iść dalej. Poniżej tego progu komórka pozostaje w stanie spoczynku. Zespół wykazał, że sztuczne aktywowanie YAP lub blokowanie górnych hamulców w szlaku Hippo konsekwentnie podwyższało stosunek cyklina D1/p27 i popychało komórki ponad ten próg, nawet w warunkach, które normalnie utrzymują komórki w ciszy.
Jak sygnały z zewnątrz są wzmacniane, a potem wygasają
YAP nie działa w pojedynkę: zwiększa wrażliwość komórki na sygnały wzrostowe pochodzące z zewnątrz. Badacze użyli sekwencjonowania RNA i pomiarów białek, aby pokazać, że YAP zwiększa liczbę i aktywność receptorów na powierzchni komórki, w tym członków rodziny EGFR i innych receptorów kinazy tyrozynowej. Receptory te zasilają klasyczne szlaki wzrostowe, takie jak MEK–ERK i, w mniejszym stopniu, mTOR, które podnoszą poziom cykliny D1 i obniżają p27, przesuwając wewnętrzny stosunek w kierunku podziału. Co istotne, to wzmocnienie nie jest szybkim włącznikiem on–off. Nawet po zablokowaniu aktywności YAP zwiększone sygnalizowanie przez receptory i podwyższony stosunek cyklina D1/p27 wygasają stopniowo przez wiele godzin, dając komórkom czas na ukończenie ograniczonej rundy podziału, zanim hamulce zostaną w pełni przywrócone.
Wbudowane zabezpieczenia przed wymknięciem się wzrostu
W badaniu przeanalizowano też, co się dzieje, gdy naturalne hamulce YAP są osłabione, jak ma to miejsce w niektórych nowotworach. Usunięcie białka łączącego o nazwie Merlin, które normalnie pomaga aktywować Hippo, częściowo uwolniło YAP i uczyniło komórki bardziej wrażliwymi na czynniki wzrostu. Nawet te komórki jednak można było wyłączyć przez bardzo wysoką lokalną gęstość komórek, co przywracało hamowanie przez kontakt i obniżało stosunek cyklina D1/p27. Pokazuje to, że system ma wielowarstwową kontrolę: receptory powierzchniowe, Hippo–YAP i stosunek cyklina D1/p27 wszystkie współdziałają przy podejmowaniu decyzji, czy komórki przystąpią do podziału.
Dlaczego to ma znaczenie dla gojenia i raka
Dla czytelnika ogólnego najważniejszy przekaz jest taki, że naprawa tkanek jest napędzana przejściowym, samograniczącym się programem wzrostu. Aktywacja YAP i zwiększone sygnalizowanie przez receptory podnoszą stosunek cyklina D1/p27 ponad próg, dzięki czemu komórki mogą się dzielić i naprawiać uszkodzenia. W miarę jak komórki się dzielą i tkanka znów staje się zatłoczona, hamowanie przez kontakt stopniowo wyłącza YAP i sygnalizowanie receptorów, stosunek spada i podziały ustają. Gdy ten mechanizm czasowy zostaje zaburzony — na przykład przez uporczywą aktywację YAP lub utratę górnych hamulców — komórki mogą stać się niewrażliwe na zagęszczenie i kontynuować podziały, co jest krokiem w kierunku nowotworu. Zrozumienie tego systemu decyzyjnego opartego na stosunku może pomóc w projektowaniu terapii regeneracyjnych, które bezpiecznie zwiększą naprawę, nie wywołując niekontrolowanego rozrostu.
Cytowanie: Ferrick, K.R., Upadhya, S.W., Fan, Y. et al. Transient proliferation by reversible YAP and mitogen control of the cyclin D1/p27 ratio. Commun Biol 9, 340 (2026). https://doi.org/10.1038/s42003-026-09590-2
Słowa kluczowe: sygnalizacja YAP, kontrola cyklu komórkowego, regeneracja tkanki, hamowanie przez kontakt, równowaga cyklina D1 p27