Clear Sky Science · pl
Droga perforująca i włókna CA3–Schaffer współdziałają, aby regulować uczenie przestrzenne
Dlaczego znajdowanie drogi ma znaczenie
Czy to mysz szukająca ukrytej platformy, czy osoba poruszająca się po nowym mieście, mózg nieustannie przetwarza wzrok, dźwięki i informacje o ruchu własnego ciała na wewnętrzną mapę. Badanie to analizuje, jak dwie kluczowe struktury mózgu — kora entorynalna i hipokamp — współpracują, by wspierać tę zdolność. Obserwując aktywność w drobnych włóknach nerwowych na żywo oraz precyzyjnie pobudzając lub wygaszając określone szlaki u myszy, badacze ujawniają, jak te obszary koordynują tworzenie i stabilizację pamięci przestrzennej — wgląd, który może w przyszłości pomóc lepiej rozumieć i leczyć zaburzenia pamięci.
Dwa kluczowe centra wewnętrznego GPS-u mózgu
Hipokamp i przylegająca kora entorynalna leżą głęboko w płacie skroniowym i tworzą rdzeń naszego systemu nawigacji. W obrębie hipokampa wewnętrzne obwody łączą obszar zwany CA3 z innym obszarem CA1 za pośrednictwem włókien znanych jako kolaterale Schaffera. Jednocześnie kora entorynalna wysyła osobny strumień informacji bezpośrednio do CA1 drogą tzw. drogi perforującej. Autorzy postanowili ustalić, jak te dwa strumienie wejściowe współdziałają, gdy zwierzę uczy się układu przestrzeni, oraz jak zmiany na ich połączeniach — ogólnie określane jako „plastyczność” — wspierają budowę stabilnej mapy przestrzennej.
Obserwowanie uczenia włókno po włóknie
Aby śledzić ten proces u zachowujących się zwierząt, zespół użył fotometrii włóknowej, techniki raportującej aktywność nerwową przez błyski fluorescencyjnego światła. Zmodyfikowali myszy tak, by neurony CA3 projecjujące do CA1 świeciły przy aktywności, a następnie trenowali zwierzęta w labiryncie wodnym Morrisa — klasycznym teście, w którym mysz musi zapamiętać ukrytą lokalizację niewielkiej platformy w basenie. W trakcie wczesnych prób włókna CA3–CA1 były silnie aktywne, gdy zwierzęta szukały; w miarę jak myszy stawały się biegłe i szybciej znajdowały platformę, ta aktywność stopniowo malała. Wzorzec sugeruje, że te połączenia są szczególnie zaangażowane podczas początkowego kodowania układu środowiska, a następnie stabilizują się w bardziej efektywną, stałą reprezentację po ustanowieniu pamięci.
Jak wejście „z góry” wzmacnia lub tłumi uczenie
Naukowcy zapytali następnie, jak sygnały z kory entorynalnej wpływają na ten obwód hipokampa. Używając światłoczułych białek do aktywacji neuronów entorynalnych podczas rejestracji z włókien CA3–CA1, wykazali, że pobudzenie wejścia z przyśrodkowej kory entorynalnej niezawodnie zwiększa aktywność tych połączeń hipokampalnych. Natomiast kiedy chemogenetycznie osłabili sygnały entorynalne docierające do CA1 podczas treningu w labiryncie wodnym, aktywność CA3–CA1 osłabła, a myszy uczyły się lokalizacji platformy wolniej i mniej precyzyjnie. W uzupełniającej serii eksperymentów autorzy rejestrowali bezpośrednio neurony entorynalne projekcjonujące do CA1 i zaobserwowali, że ich aktywność narastała w kolejnych dniach treningu, odzwierciedlając poprawę wyników zwierząt. Razem wyniki te wskazują, że silne, dobrze zgrane sygnały entorynalne są potrzebne do odpowiedniego dostrojenia obwodów hipokampa pod kątem nawigacji.
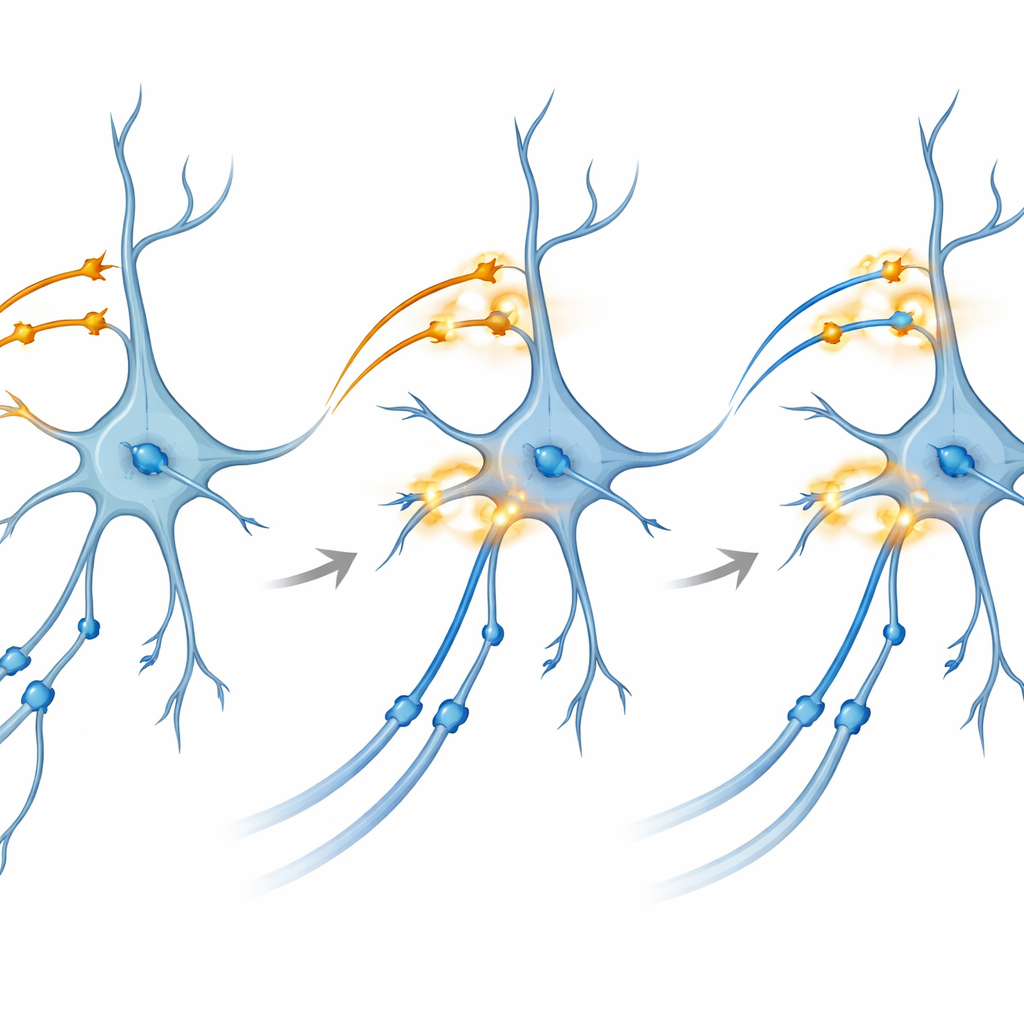
Zbliżenie na mechanizm wzmacniania połączeń
Aby zbadać mechanizm leżący u podstaw, zespół sięgnął po plasterki mózgu utrzymywane przy życiu in vitro. Tam mogli oddzielnie kontrolować światłoczułe białka w CA3 i w wejściach entorynalnych do CA1. Ku zaskoczeniu, stymulacja błyskowa w rytmie theta — wzory naśladujące naturalne rytmiczne wyładowania — zastosowana osobno na którymkolwiek szlaku nie wywoływała niezawodnie długotrwałego wzmocnienia (LTP), trwałego wzmocnienia synaps uważanego za podstawę pamięci. Jednak gdy dostarczyli starannie zsynchronizowany dwukolorowy impuls, który jednocześnie współaktywował oba szlaki do CA1, połączenia CA3–CA1 wykazały silne i długotrwałe wzmocnienie. Zablokowanie receptorów NMDA lub specyficznych kanałów wapniowych — dwóch dobrze znanych „bram” molekularnych plastyczności — uniemożliwiło ten efekt, łącząc zjawisko z klasycznymi szlakami biochemicznymi tworzenia pamięci.
Co to oznacza dla pamięci i chorób
W całości badanie przedstawia obraz uczenia się przestrzennego jako skoordynowanego tańca między napływającymi sygnałami entorynalnymi a wewnętrznym okablowaniem hipokampa. Wejścia entorynalne nie tylko przenoszą informacje o pozycji i kontekście; działają też jako potężny regulator, który może wzmacniać lub tłumić aktywność włókien CA3–CA1 i, gdy są współaktywowane, napędzać długotrwałe wzmocnienie tych synaps. Dla czytelnika popularnonaukowego kluczowy wniosek jest taki, że „GPS” mózgu nie opiera się na jednym obszarze, lecz na partnerstwie, w którym jeden region ustawia scenę, a drugi utrwala szczegóły. Zakłócenia tego partnerstwa — spowodowane wiekiem, urazem czy chorobą neurodegeneracyjną — mogą więc pomóc wyjaśnić, dlaczego ludzie gubią się w znajomych miejscach, a ukierunkowanie tych szlaków może otworzyć nowe drogi zachowania lub przywracania pamięci przestrzennej.
Cytowanie: Huang, F., Temitayo Bello, S., Lau, S.H. et al. The perforant pathway and CA3-Schaffer collateral afferents coordinate to regulate spatial learning. Commun Biol 9, 364 (2026). https://doi.org/10.1038/s42003-026-09577-z
Słowa kluczowe: pamięć przestrzenna, hipokamp, kora entorynalna, plastyczność synaptyczna, nawigacja