Clear Sky Science · pl
Interkortykalny interfejs mózg–maszyna oparty na aktywności brzusznej kory przedruchowej makaka
Nauczanie mózgu poruszania kursorem
Wyobraź sobie sterowanie kursorem komputera lub ramieniem robota jedynie myślami, nawet jeśli twoje mięśnie nie mogą się poruszyć. Interfejsy mózg–maszyna (BMI) mają to umożliwić poprzez tłumaczenie aktywności mózgu na polecenia dla urządzeń zewnętrznych. Większość dotychczasowych systemów korzystała z jednego głównego obszaru ruchowego mózgu — co jednak, gdy ten region jest uszkodzony, na przykład w wyniku udaru lub stwardnienia zanikowego bocznego? W tym badaniu postawiono pytanie, czy inny, sąsiadujący obszar, zwykle kojarzony z planowaniem ruchów ręki i obserwowaniem ruchów innych, również może niezawodnie napędzać BMI.
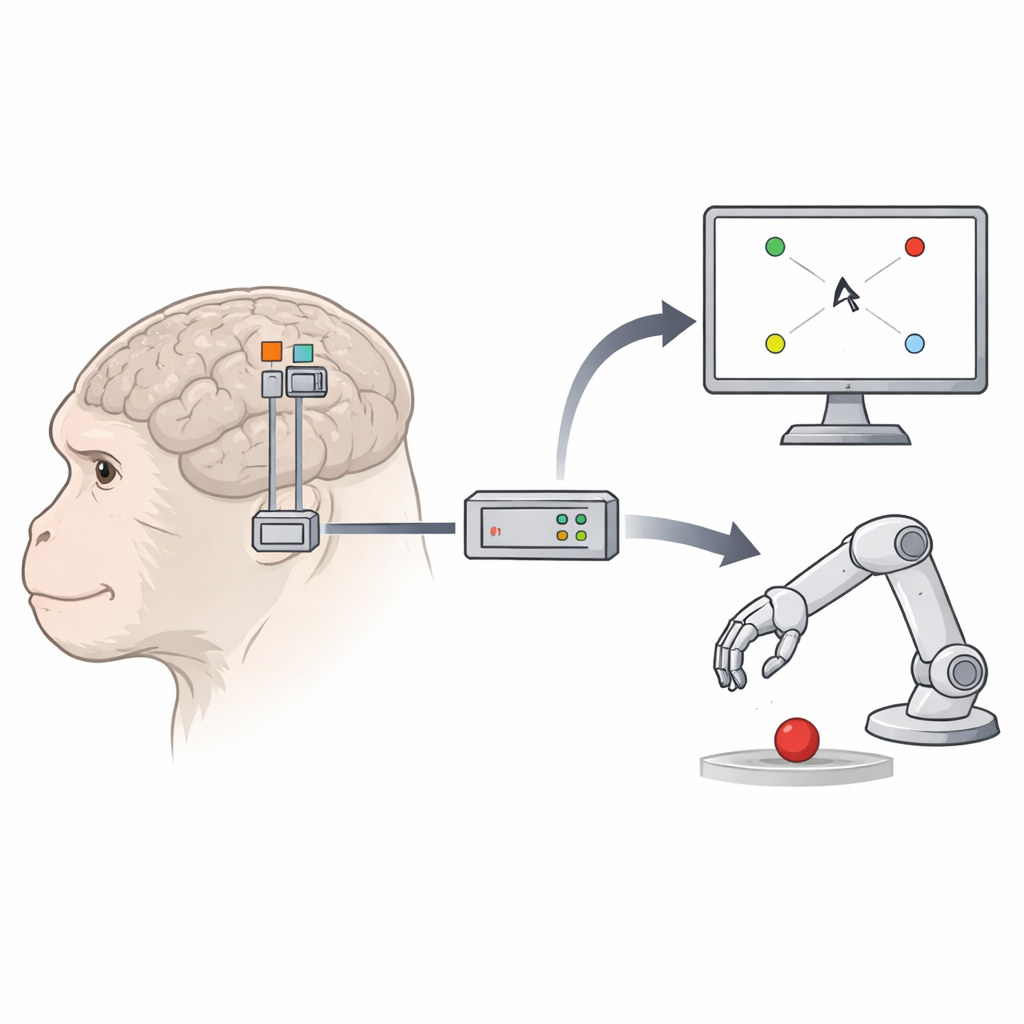
Nowy obszar dołącza do zespołu
Klasyczne BMI odczytują głównie sygnały z pierwotnej kory ruchowej — paska tkanki mózgowej, który bezpośrednio kontroluje ruchy dowolne — oraz z przyległego obszaru planowania zwanego grzbietową korą przedruchową. Badacze zwrócili uwagę na innego sąsiada: brzuszną korę przedruchową, a konkretnie strefę nazwaną F5c. U małp F5c jest bogata w komórki, które aktywują się, gdy zwierzę sięga i chwyta przedmioty, a nawet gdy jedynie obserwuje akcje na ekranie. To połączenie reakcji związanych zarówno z wykonywaniem ruchu, jak i jego obserwacją sugerowało, że F5c może być dobrze przystosowana do sterowania kursorem lub „awatarem” robota bez konieczności poruszania ciałem.
Małpy, mikroelektrody i cele ruchome
Dwie makaki otrzymały implanty z drobnymi siatkami 96 elektrod w trzech miejscach: w pierwotnej korze ruchowej, grzbietowej korze przedruchowej i F5c. Podczas codziennych sesji zwierzęta wykonywały kilka wizualnie prostych, lecz behawioralnie wymagających zadań. W jednym zadaniu dotykały środka ekranu, a następnie sięgały do jednego z ośmiu zewnętrznych celów, podczas gdy mały kwadratowy kursor na ekranie przesuwał się ze środka do tego samego celu. W drugim zadaniu trzymały rękę nieruchomo i jedynie obserwowały ruch kursora do celów. W trzeciej, bardziej realistycznej konfiguracji, kursor został zastąpiony awatarem 3D — ramieniem robota sięgającym do celów w wirtualnej scenie. W tych zadaniach zespół mógł porównać, jak dobrze każdy obszar mózgu napędza ruch kursora lub awatara.
Jak sygnały mózgowe zamieniły się w płynny ruch
W fazie treningowej kursor lub awatar podążały po komputerowo wygenerowanych, łagodnie zakrzywionych ścieżkach, podczas gdy małpy albo się poruszały, albo obserwowały. Jednocześnie elektrody rejestrowały szybkie wybuchy aktywności mózgowej. Następnie badacze wytrenowali dekoder — narzędzie matematyczne uczące się odwzorowywać wzorce strzałów neuronalnych na prędkości kursora lub awatara na ekranie. Aby wybrać tylko najbardziej informatywne kanały, użyli elektrod, których aktywność śledziła kierunek i prędkość ruchu. Zastosowali metodę wyodrębniającą wzorce mózgowe najściślej powiązane z zachowaniem i wzbogacili ją krokiem nieliniowym, co pozwoliło systemowi uchwycić bardziej złożone relacje między aktywnością neuronalną a ruchem. W fazie dekodowania komputer przestał sterować kursorem lub awatarem; zamiast tego dekoder używał na żywo sygnałów mózgowych, aktualizowanych co 50 milisekund, by kierować ruchem na ekranie. Dekoder był okresowo ponownie trenowany w tle, aby móc się adaptować, gdy odpowiedzi neuronów zmieniały się w czasie.

Jak poradził sobie „nowy” obszar?
Kluczowe pytanie brzmiało, czy F5c może dorównać lub znaleźć się blisko wydajności tradycyjnych obszarów kontrolnych. U obu małp dekodowanie oparte na F5c na początku ustępowało, zwłaszcza gdy poruszano kursorem, a zwierzęta jedynie obserwowały i nie ruszały własną ręką. Jednak w miarę postępu sesji — i gdy więcej elektrod dostarczało wiarygodnych sygnałów związanych z ruchem — F5c doganiało resztę. W kilku warunkach jego wydajność była równa pierwotnej korze ruchowej, a nawet ją przewyższała w późniejszych sesjach przy pasywnym sterowaniu kursorem. Przy sterowaniu awatarem robota ogólny współczynnik sukcesu był niższy we wszystkich obszarach, lecz F5c nadal wspierało znaczące sterowanie, zwłaszcza w połączeniu z łagodnym algorytmem wspomagającym, który subtelnie kierował awatara w stronę celu. Co ważne, neurony we wszystkich trzech regionach wykazywały podobne wzorce strojenia kierunkowego i aktywności populacyjnej podczas fazy treningowej sterowanej komputerowo i fazy aktywnego sterowania, przy czym jedynie podzbiór zmieniał preferowane kierunki w miarę jak małpy uczyły się prowadzić BMI.
Co to oznacza dla przyszłej neurotechnologii
Dla osoby niebędącej specjalistą kluczowy wniosek jest taki, że obszar brzusznej kory przedruchowej F5c — wcześniej postrzegany głównie jako centrum planowania i obserwacji działań — może również służyć jako praktyczne centrum sterowania dla interfejsów mózg–maszyna. Gdy dostępnych jest wystarczająco dużo sygnałów związanych z ruchem, dekodery wytrenowane na aktywności F5c mogą prowadzić kursor na ekranie lub wspomagać robota niemal tak samo dobrze jak te oparte na klasycznym obszarze ruchowym. Sugeruje to, że przyszłe kliniczne BMI nie będą musiały polegać na jednym korowym regionie. Dla osób z uszkodzoną pierwotną korą ruchową pobliskie obszary planowania, takie jak brzuszna kora przedruchowa, mogą stanowić alternatywną drogę do odzyskania kontroli nad narzędziami cyfrowymi, urządzeniami protetycznymi czy pomocami mobilności.
Cytowanie: De Schrijver, S., Garcia Ramirez, J., Iregui, S. et al. An intracortical brain-machine interface based on macaque ventral premotor activity. Sci Rep 16, 8407 (2026). https://doi.org/10.1038/s41598-026-38536-1
Słowa kluczowe: interfejs mózg–maszyna, kora ruchowa, kora przedruchowa, dekodowanie neuronalne, sterowanie protezą