Clear Sky Science · pl
Przestrzenno-czasowa ekspresja przyczepów endospor i wgląd z cryo-EM w kotwiczenie S-ENA przez Ena1C w Bacillus paranthracis
Dlaczego uparte spory bakteryjne mają znaczenie
Bakterie z rodziny Bacillus cereus mogą przekształcać się w wytrzymałe spory, które są odporne na działanie wysokiej temperatury, substancji chemicznych i środków czyszczących. Te spory często przylegają do urządzeń przetwórstwa żywności i innych powierzchni, gdzie mogą wywoływać zatrucia pokarmowe lub długo zanieczyszczać produkty. W tym badaniu przyglądamy się drobnym włóknistym wypustkom na tych sporach, zwanym przyczepami endospor (endospore appendages), i ujawniamy, kiedy są one budowane oraz jak kluczowe białko Ena1C mocuje je do powierzchni spory. Zrozumienie tego procesu może pomóc przemysłowi w projektowaniu skuteczniejszych strategii czyszczenia, a także zainspirować nowe sposoby inżynierii użytecznych spor do zastosowań rolniczych lub biotechnologicznych.
Sztywne włoski na uśpionych bakteriach
W mikroskopie spory wielu bakterii z grupy Bacillus cereus pokryte są cienkimi włóknami przypominającymi włoski lub pilusy. Te przyczepy endospor są wyjątkowo odporne i opierają się zarówno enzymom, jak i silnym środkom chemicznym, co utrudnia ich badanie. Wcześniejsze prace przy użyciu zaawansowanej krio-elektronowej mikroskopii ujawniły, że Bacillus paranthracis wytwarza dwa główne typy włókien: grube, przestawione S-ENA, które stanowią około 90% włosków, oraz cieńsze, przypominające drabinkę L-ENA, które stanowią resztę. Analizy genetyczne powiązały już konkretne geny ena z każdym typem włókien, jednak kiedy i gdzie te białka pojawiają się podczas formowania spory oraz jak S-ENA są rzeczywiście zakotwiczone do spory, pozostawały otwartymi pytaniami.
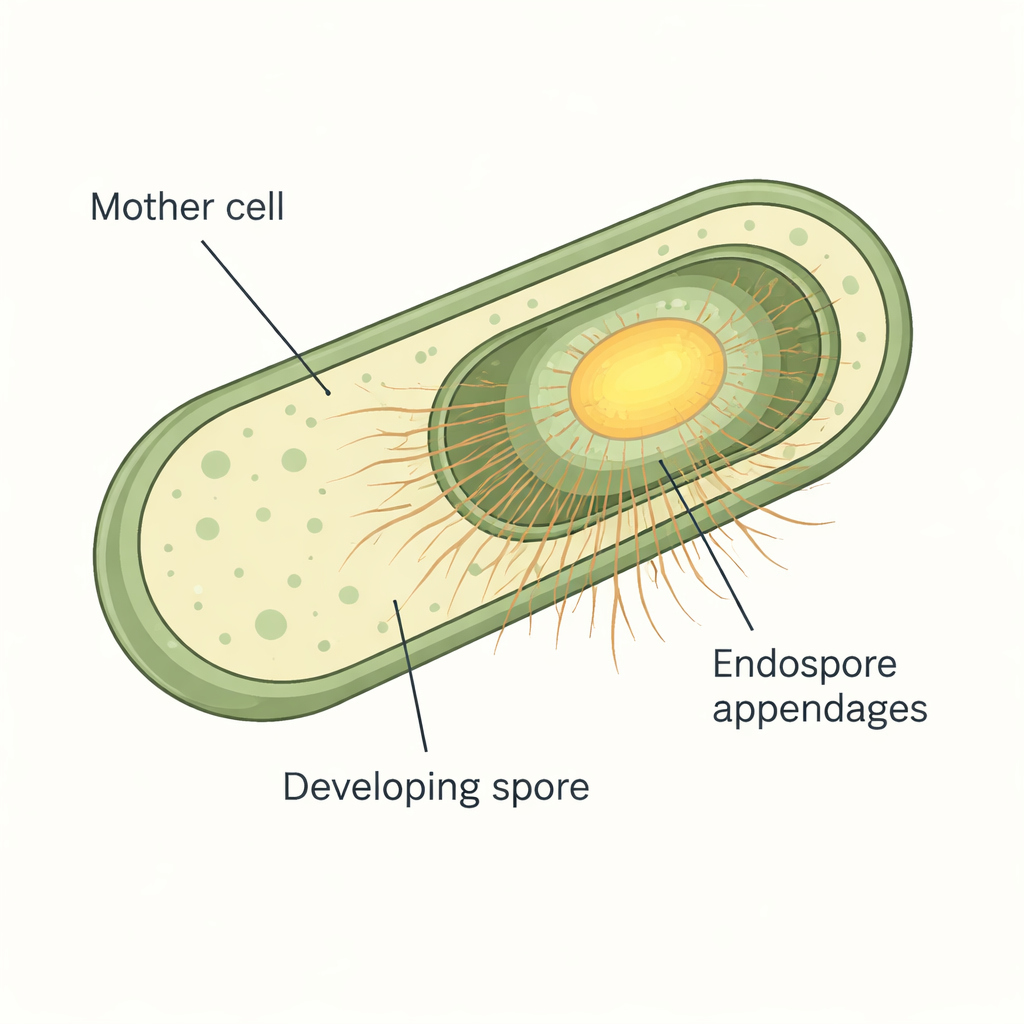
Obserwowanie włókien włączających się w czasie rzeczywistym
Aby śledzić te elementy budulcowe podczas formowania spor, badacze połączyli białka ENA z jasnymi znacznikami fluorescencyjnymi, co pozwoliło im obserwować każde białko w żywych komórkach w mikroskopii w czasie. Hodowali Bacillus paranthracis na specjalnie przygotowanych podłożach agarowych pod mikroskopem i rejestrowali obrazy co 10–12 minut, podczas gdy komórki przechodziły przez sporulację. Zespół stwierdził, że żadne z białek ENA nie były produkowane podczas normalnego wzrostu komórek. Produkcja rozpoczynała się dopiero po tym, jak rozwijające się spory stały się „faza-jasne” (phase-bright) — wizualnym sygnałem, że rdzeń spory i warstwy ochronne dojrzały. Fluorescencja ENA następnie gwałtownie rosła późno w sporulacji i koncentrowała się wokół spory, szczególnie na styku między komórką macierzystą a formującą się sporą, co świadczy, że te włókna są rzeczywiście dekoracjami specyficznymi dla spory dodawanymi pod koniec procesu.
Dwa typy włókien w różnych harmonogramach
Zespół porównał także opóźnienia w produkcji S-ENA i L-ENA, śledząc jednocześnie dwa białka w tych samych komórkach. Gdy podjednostki S-ENA (Ena1A lub Ena1C) oznaczone na zielono były koekspresjonowane z podjednostką L-ENA Ena3A oznaczoną na czerwono, sygnał zielony pojawiał się konsekwentnie około godzinę przed czerwonym. Ten przesunięty harmonogram odpowiada temu, co wiadomo o architekturze spory: S-ENA wyłaniają się z płaszcza spory, który formuje się wcześniej, podczas gdy L-ENA są zakotwiczone w egzosporium, zewnętrznym „woreczku” dodawanym później. Wyniki sugerują, że komórka stosuje późne przełączniki genetyczne, aby włączać geny ENA w ścisłej kolejności, tak by każdy typ włókien trafił do właściwej warstwy spory we właściwym czasie.
Molekularny „dok”, który trzyma włókna na miejscu
Jednym z najbardziej zagadkowych uczestników był Ena1C — białko niezbędne do pojawienia się włókien S-ENA na sporach, choć nie będące częścią samego trzonu włókna. Analiza spor pochodzących od bakterii pozbawionych genu ena1C wykazała, że włókna S-ENA nadal się składały, lecz unosiły się swobodnie w otaczającej cieczy zamiast być przymocowane do spor. Kiedy Ena1C było nadmiernie produkowane, spory nosiły znacznie więcej S-ENA, ale każde włókno było krótsze, jakby ograniczona pula elementów budulcowych była rozdzielana na większą liczbę punktów przyczepu. To wskazywało na Ena1C jako dedykowaną kotwicę, która przywiązuje włókna S-ENA do płaszcza spory, kontrolując zarówno liczbę przyczepionych włókien, jak i ich długość.
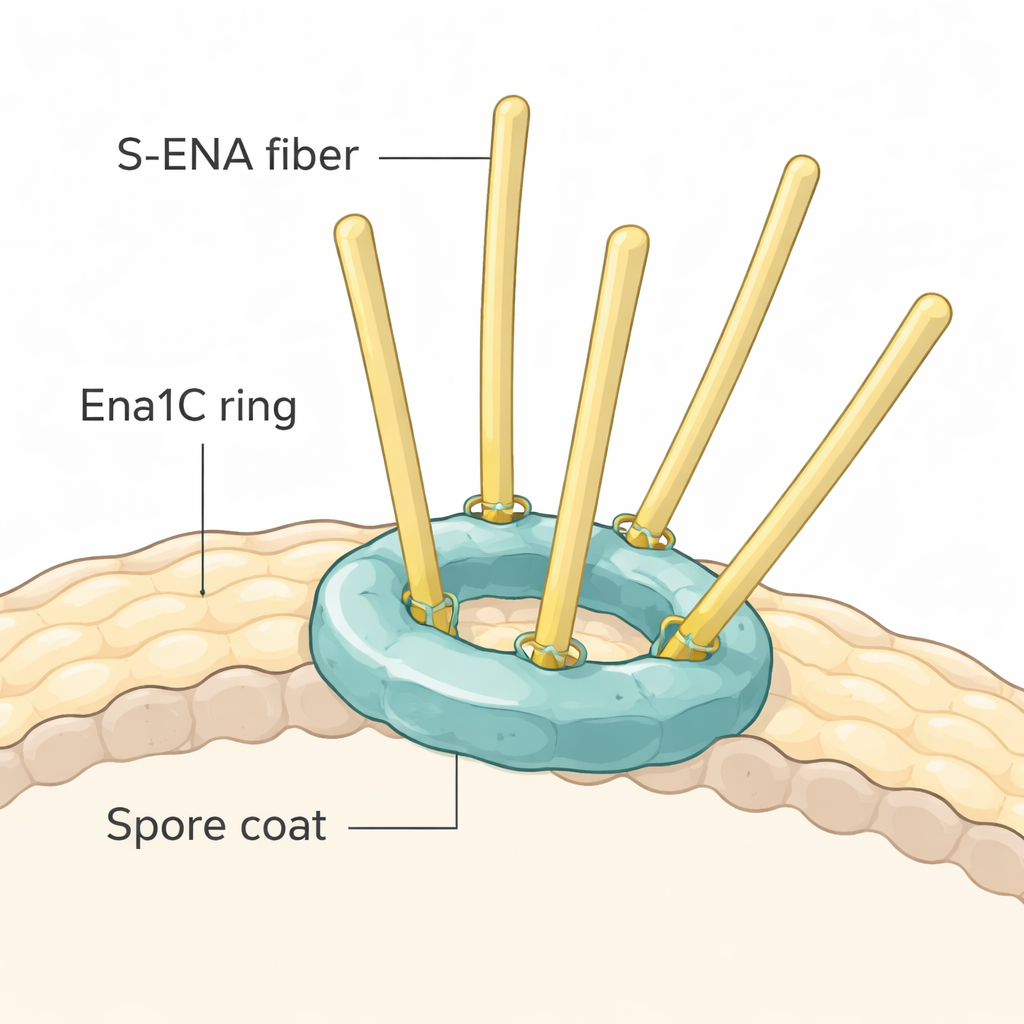
Odkrycie kotwic w kształcie pierścienia dzięki cryo-EM
Aby zobaczyć, jak Ena1C może chwytać włókna, badacze oczyścili to białko i obrazowali je przy użyciu wysokorozdzielczej krio-elektronowej mikroskopii. Odkryli, że Ena1C nie tworzy długich włókien. Zamiast tego dziewięć kopii białka łączy się, tworząc solidny pierścień z centralnym otworem, wzmocniony licznymi mostkami disiarczkowymi — silnymi wiązaniami chemicznymi między aminokwasami zawierającymi siarkę. Modelowanie komputerowe i porównania strukturalne z znanymi białkami włóknistymi sugerują, że trzon S-ENA prawdopodobnie zadokowuje w dodatnio naładowanym lejku u środka tego pierścienia, gdzie kluczowe reszty cysteinowe Ena1C mogą tworzyć mostki disiarczkowe z odpowiadającymi miejscami na podjednostkach włókna. W ten sposób każdy pierścień Ena1C wydaje się działać jak molekularny dok, który zaciska jedno lub kilka włókien S-ENA do zewnętrznego płaszcza spory.
Co to oznacza dla kontroli uparcie trzymających się spor
Podsumowując, badanie pokazuje, że włoski spor Bacillus paranthracis są produkowane dopiero po osiągnięciu dojrzałości spory, przy czym włókna S-ENA są budowane i kotwiczone do płaszcza przed dodaniem L-ENA do egzosporium. Identyfikuje także Ena1C jako pierścieniowatą kotwicę, która kowalencyjnie wiąże włókna S-ENA z powierzchnią spory. Dla laików kluczowy wniosek jest taki, że sposób, w jaki spory przylegają do powierzchni, nie jest przypadkowy: to efekt starannie zaplanowanej budowy, która dodaje wytrzymałe, włókniste „rzepy” na zewnątrz spory. Celując w ten system kotwiczenia — poprzez zakłócenie Ena1C lub zmianę tworzenia ENA — przyszłe strategie mogą osłabić przyleganie spor, ułatwiając czyszczenie przemysłowe, lub przeciwnie, pozwolić inżynierom zaprojektować spory, które celowo przyczepiają się do upraw lub materiałów w korzystny sposób.
Cytowanie: Zegeye, E.D., Sleutel, M., Jonsmoen, U.L. et al. Spatiotemporal expression of endospore appendages and cryo-EM insights into Ena1C-mediated S-ENA anchoring in Bacillus paranthracis. Sci Rep 16, 7122 (2026). https://doi.org/10.1038/s41598-026-38321-0
Słowa kluczowe: spory bakteryjne, przyleganie powierzchniowe, mikroskopia krioelektronowa, składanie białek, bezpieczeństwo żywności