Clear Sky Science · pl
Sprzężenie hipokamp–jądro ponadmammilowe w przebiegu snu i czuwania
Dlaczego to ma znaczenie dla codziennej pamięci
Co noc, gdy śpimy, a nawet gdy w spokoju odpoczywamy, nasze mózgi odtwarzają doświadczenia i przekształcają wspomnienia. Badanie to przygląda się, jak kluczowe centrum pamięciowe — hipokamp — komunikuje się z głębszymi ośrodkami mózgu, które kontrolują pobudzenie, ruch i nastrój. Obserwując te regiony podczas różnych stanów snu i czuwania u szczurów, autorzy ujawniają, jak mózg elastycznie przekierowuje informacje — czasem wysyłając silne sygnały „z góry” z obwodów pamięciowych, innym razem pozwalając głębszym rejonom chwilowo przejąć inicjatywę. Zrozumienie tej wymiany pomaga wyjaśnić, jak sen wspiera pamięć, jednocześnie utrzymując stabilność podstawowych funkcji organizmu.
Dwukierunkowe rozmowy między ośrodkami pamięci i pobudzenia
Zespół rejestrował drobne sygnały elektryczne z hipokampa oraz z dwóch połączonych rejonów — jądra ponadmammilowego i przegrody bocznej — u swobodnie poruszających się szczurów. Śledzili zwierzęta w czasie cichego odpoczynku, głębokiego snu NREM (bez szybkich ruchów gałek ocznych), snu REM bogatego w marzenia oraz aktywnego czuwania. Wykorzystując te zapisy, śledzili krótkie wybuchy zsynchronizowanej aktywności hipokampa, zwane ripple’ami, i związane z nimi zdarzenia w pobliskim rejonie zwanym gąbczastym zakrętem (dentate gyrus). Równocześnie mierzyli częstość wyładowań neuronów w jądrze ponadmammilowym, które wpływa na pobudzenie i stadia snu, oraz w przegrodzie bocznej łączącej obwody pamięciowe z systemami motywacji i ruchu.
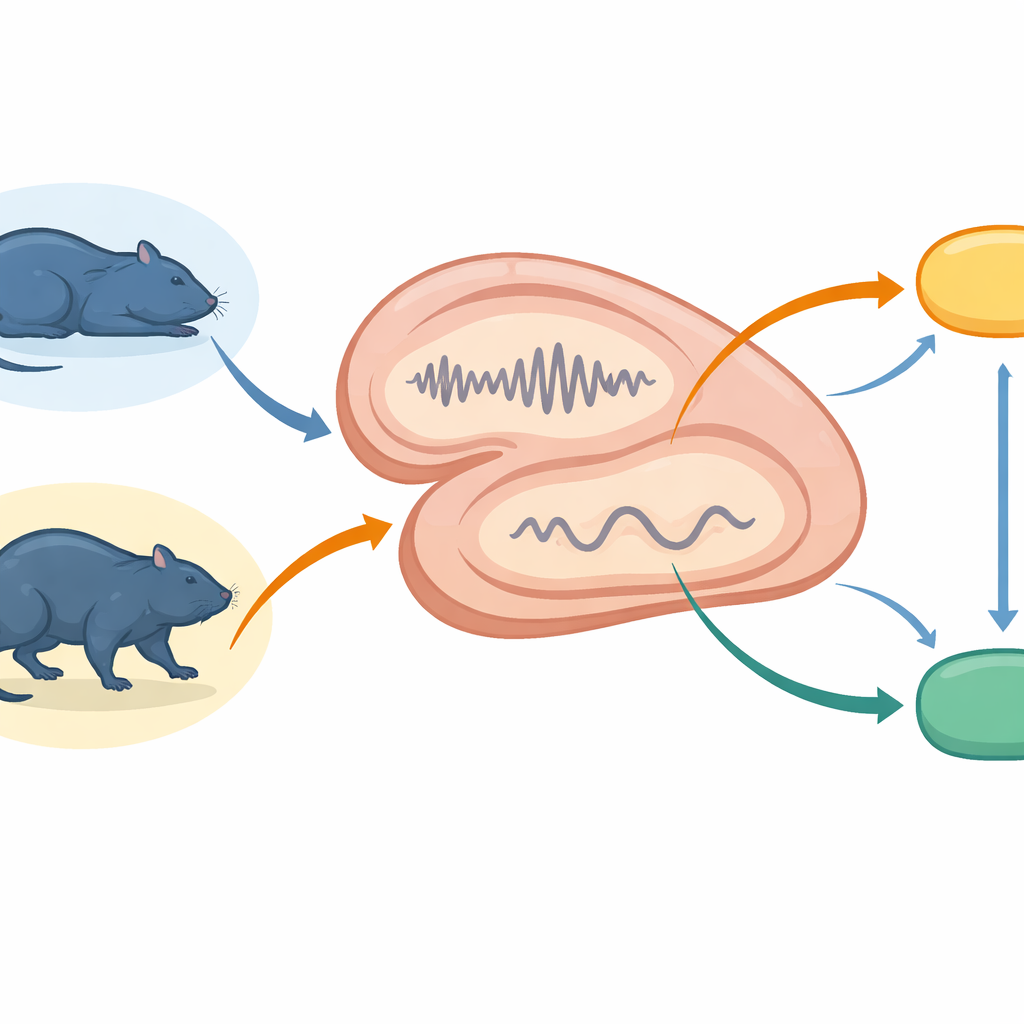
Impulsy „z góry” w cichych momentach
Podczas cichego czuwania i snu NREM — stanów sprzyjających odtwarzaniu pamięci — hipokamp często generował ostre, szybkie zdarzenia typu ripple. Ripple wywoływały duże surżenia wyładowań lokalnie w hipokampie oraz mniejsze, lecz powtarzalne aktywacje w jądrze ponadmammilowym i przegrodzie bocznej. Innymi słowy, system pamięci nadawał krótkie impulsy „z góry” do głębszych ośrodków, ale o mniejszej sile niż w obrębie własnej aktywności. Zdarzenia typu dentate spike, inny rodzaj szybkich epizodów w pobliskim podregionie hipokampa, powodowały słabsze i bardziej jednorodne reakcje, które ledwo docierały do obszarów podkorowych. Ten kontrast sugeruje, że ripple są głównym nośnikiem szybkiej, szeroko zakrojonej koordynacji, podczas gdy dentate spike dopracowują lokalne przetwarzanie bez silnego pobudzania dalszych centrów.
Impulsy „z dołu” od wybuchów głębokiego mózgu
Komunikacja nie była jednokierunkowa. Kiedy jądro ponadmammilowe wchodziło w krótkie, wysokiej aktywności epizody, wysyłało sygnały „z dołu” z powrotem do hipokampa. Miało to bardzo różne konsekwencje zależnie od stanu mózgu. Podczas snu NREM takie wybuchy chwilowo tłumiły wyładowania w kluczowym obszarze wyjściowym hipokampa (CA1), podczas gdy neurony gąbczastego zakrętu zwiększały aktywność wolniej. W cichym czuwaniu te same wybuchy jądra ponadmammilowego powodowały słabsze zahamowanie CA1, ale wyraźniejszy netto wzrost ogólnej aktywności hipokampa. Wzorce te wskazują, że wybuchy głębokiego mózgu mogą albo tłumić, albo wzmacniać wyjście hipokampa, regulując, jak silnie obwody pamięciowe wpływają na resztę mózgu, bez ich całkowitego wyłączania.
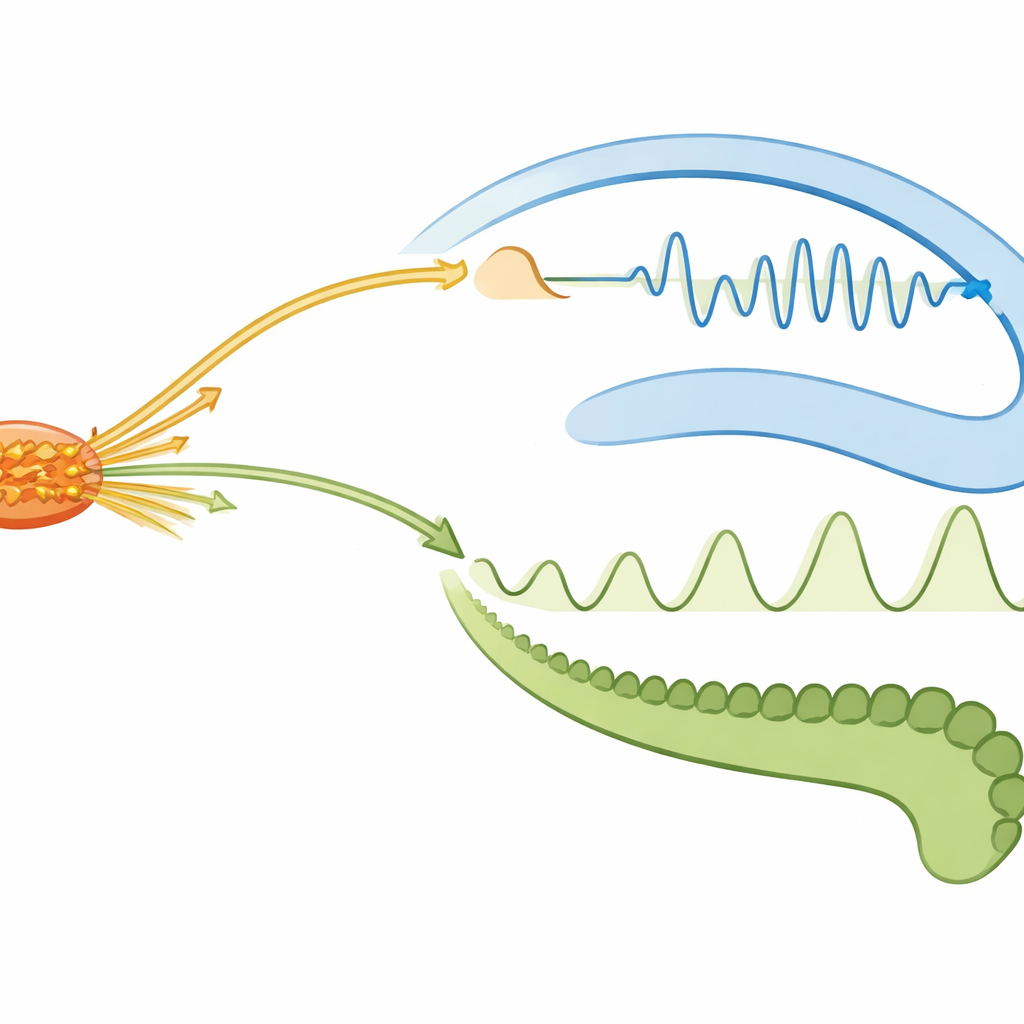
Rytmiczne timingowanie podczas aktywnych i śniących stanów
Gdy mózg przechodził w stany bogate w falę theta — aktywne eksplorowanie podczas czuwania i żywe marzenia podczas snu REM — dialog znów się zmieniał. Fale theta to wolniejsze rytmy, które pomagają koordynować wyładowania między regionami. W aktywnym czuwaniu theta organizowała powtarzalny porządek wystrzałów: neurony gąbczastego zakrętu miały tendencję do wyładowań jako pierwsze w każdej cykli, potem przegroda boczna, a następnie CA1 w pobliżu dołka fali, podczas gdy jądro ponadmammilowe wykazywało mało spójnego timingowania. W tym stanie wybuchy jądra ponadmammilowego wywoływały szybkie, krótkotrwałe zahamowanie w CA1, pozostawiając aktywność gąbczastego zakrętu w dużej mierze nienaruszoną — co sugeruje selektywne „bramkowanie” niektórych wejść do hipokampa. W śnie REM układ się odwracał: neurony jądra ponadmammilowego silnie synchronizowały się z rytmem theta, wyładowując blisko szczytu każdego cyklu, podczas gdy komórki hipokampa i przegrody miały słabsze preferencje fazowe. Ta zmiana sugeruje, że podczas marzeń głębokie ośrodki pobudzeniowe przyjmują bardziej rytmiczną, koordynującą rolę.
Co to oznacza dla snu, pamięci i równowagi mózgu
Wszystko razem wyniki ujawniają elastyczne, zależne od stanu partnerstwo między hipokampem a jego podkorowymi partnerami. W stanach cichych ostre ripple pozwalają hipokampowi wysyłać potężne, ale starannie ograniczone impulsy na zewnątrz, prawdopodobnie wspierając odtwarzanie pamięci bez wywoływania dużych zmian w pobudzeniu czy zachowaniu. W bardziej aktywnych stanach zdominowanych przez theta krótkie wybuchy jądra ponadmammilowego mogą selektywnie tłumić lub timingować wyjście hipokampa, szczególnie w CA1, podczas gdy sen REM pozwala tym głębokim neuronom rytmicznie synchronizować się z obwodami pamięciowymi. Dla czytelnika nietechnicznego kluczowa myśl jest taka, że regiony pamięci i ośrodki pobudzenia prowadzą nieustanną, dwukierunkową negocjację, dostosowując kto przewodzi, a kto podąża w miarę przejścia od czuwania do różnych stadiów snu — co pomaga zachować wspomnienia przy jednoczesnym utrzymaniu ogólnej stabilności mózgu.
Cytowanie: Caneo, M., Espinosa, N., Lazcano, G. et al. Hippocampo–supramammillary coupling across sleep and wake. Sci Rep 16, 10465 (2026). https://doi.org/10.1038/s41598-026-37066-0
Słowa kluczowe: hipokamp, sen i pamięć, rytmy mózgowe, jądro ponadmammilowe, komunikacja neuronalna