Clear Sky Science · pl
Sororin blokuje bramkę wyjścia DNA w kohezynie, aby zachować spójność chromatyd siostrzanych
Utrzymywanie skopiowanych chromosomów razem
Za każdym razem, gdy komórka się dzieli, musi skopiować swoje DNA i przekazać pełny zestaw każdej komórce potomnej. Jeśli te kopie rozdzielą się za wcześnie, efektem mogą być pęknięte chromosomy, genetyczny chaos i choroby, w tym rak. W tym badaniu odkryto, jak mały fragment białka zwany ogonem Sororin działa niczym fizyczna zasuwka na pierścieniowym zacisku, który utrzymuje razem siostrzane kopie DNA, wyjaśniając, jak komórki parują chromosomy aż do dokładnego momentu rozdzielenia.
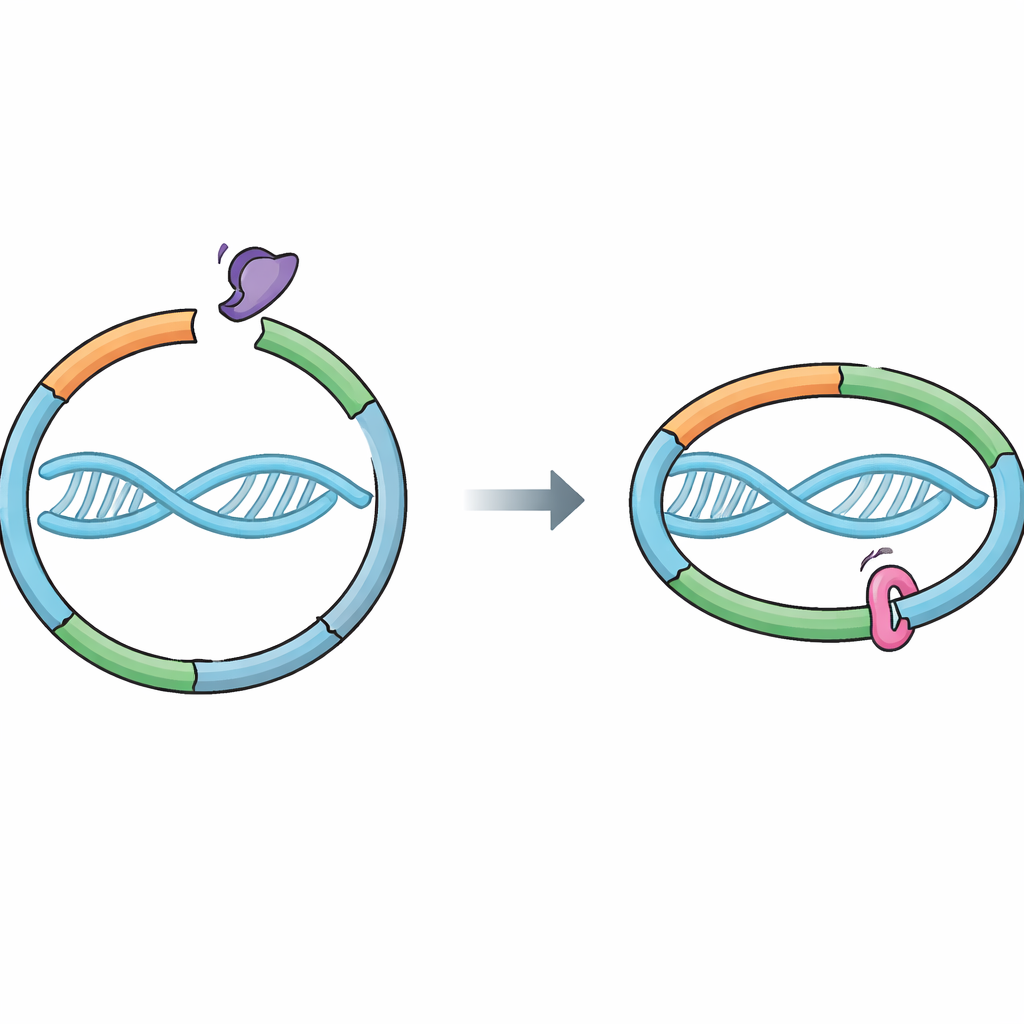
Molekularny pierścień strzegący naszych genów
Komórki polegają na maszynie białkowej zwanej kohezyną, która trzyma razem świeżo skopiowane chromosomy. Kohezyna tworzy pierścień, który oplata DNA, łącząc dwie siostrzane nici utworzone podczas replikacji. To połączenie musi być wystarczająco mocne, by przetrwać wszystkie wstrząsy podziału komórkowego, a jednocześnie podatne na rozluźnienie w odpowiednim momencie, aby siostry mogły się rozdzielić czysto. Białko pomocnicze Wapl sprzyja otwieraniu specyficznej „bramki wyjścia” w pierścieniu w miejscu spotkania dwóch części kohezyny, RAD21 i SMC3. Otwarcie tej bramki pozwala DNA wysunąć się i kohezynie odpaść. Do tej pory sądzono, że Sororin zachowuje spójność głównie przez blokowanie partnera Wapl, Pds5, ale nie było jasne, czy Sororin także bezpośrednio dotyka pierścienia kohezyny.
Maleńki ogon z wielką siłą trzymania
Autorzy skupili się na skrajnym końcu ogona Sororin — ostatnich 30 aminokwasach, zwanych regionem C‑terminalnym (CTR). Poprzez przycinanie Sororin na fragmenty i testy w komórkach ludzkich stwierdzili, że ten drobny CTR jest zarówno konieczny, jak i wystarczający do utrzymania siostrzanych chromatyd razem. Gdy CTR został sztucznie umieszczony przy centromerach, centralnych miejscach, gdzie siostry pozostają najdłużej złączone, prawie całkowicie przywrócił spójność w komórkach pozbawionych pełnego Sororin. Odwrotnie, gdy CTR był przytwierdzony wzdłuż całego chromatyny, kohezyna nie mogła być skutecznie usunięta, a chromosomy nie kompaktowały się i nie rozplątywały prawidłowo. Te komórki miały długie, rozmyte chromosomy mitotyczne, mostki DNA między rozdzielającymi się siostrami i błędy segregacji — problemy przypominające te, które występują po usunięciu samego Wapl.
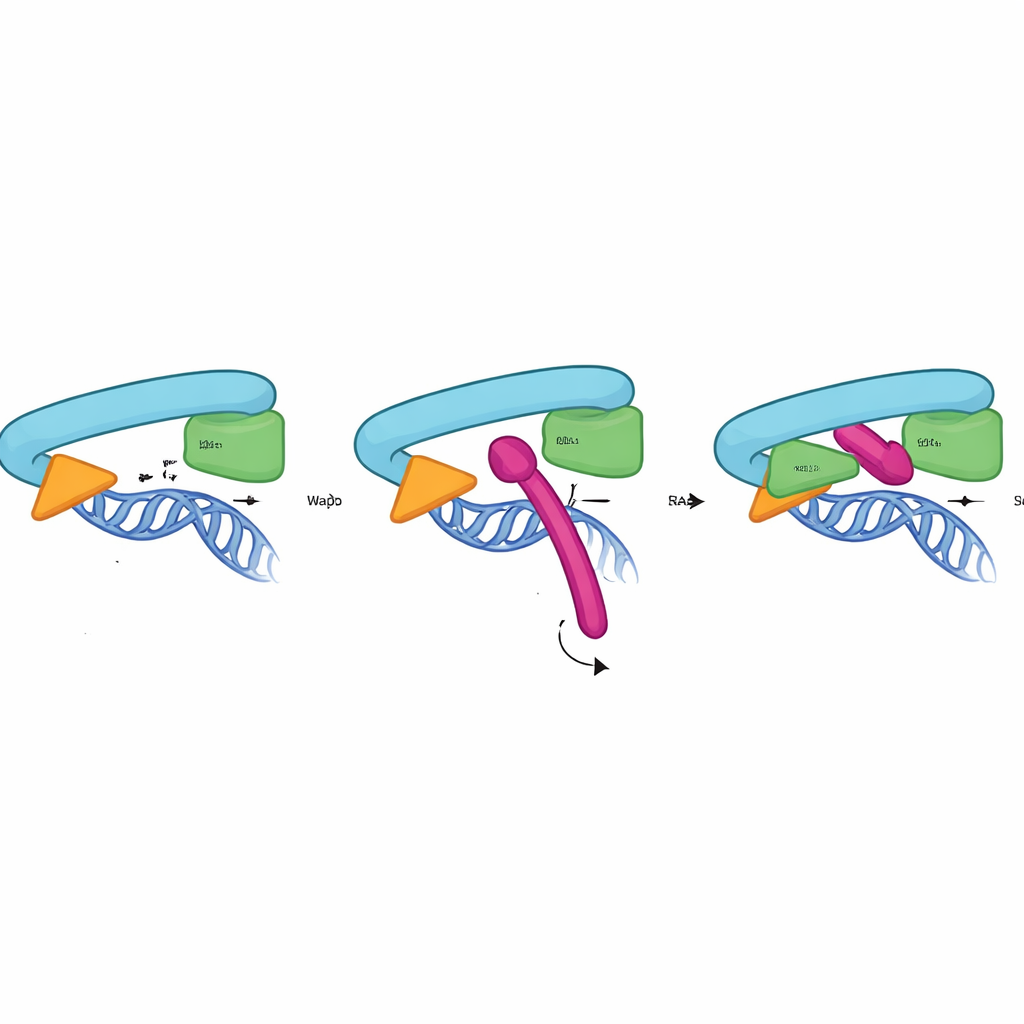
Jak zamek Sororin uchwytuje pierścień
Aby zrozumieć działanie CTR na poziomie atomowym, badacze połączyli testy biochemiczne, mutagenezę i predykcje struktur AlphaFold3. Wykazali, że CTR wiąże się bezpośrednio i specyficznie z interfejsem między RAD21 a głową SMC3 — dokładnie w miejscu tworzącym bramkę wyjścia DNA. To oddziaływanie nie wymaga Pds5 ani podjednostki SA2 i zachodzi tylko na kohezynie związanej z chromatyną, osiągając szczyt w fazie, gdy spójność musi być stabilnie utrzymana. Model strukturalny sugeruje, że krótka helisa w obrębie CTR wsuwa się w kieszeń utworzoną przez RAD21 i SMC3, wykorzystując skupisko zachowanych hydrofobowych i ujemnie naładowanych reszt, by się zaczepić. Zmutowanie zaledwie kilku z tych punktów kontaktowych w Sororinie, RAD21 lub SMC3 wystarczyło, by zniweczyć wiązanie i spowodować przedwczesne rozdzielenie sióstr, co potwierdza, że ten interfejs działa jako fizyczna zasuwka bramki.
Wyłącznik czasowy do rozdziału chromosomów
Komórki muszą też uwolnić kohezynę w odpowiednim momencie. Zespół odkrył, że Sororin przenosi pobliskie miejsce kontroli — pojedynczy aminokwas (S145), który ulega fosforylacji przez kinazę mitotyczną Aurora B. Ta modyfikacja specyficznie przerywa uchwyt Sororin za Pds5, pozwalając Wapl ponownie działać i promować usuwanie kohezyny z ramion chromosomów, ale pozostawia kontakt CTR z bramką RAD21–SMC3 w dużej mierze nietknięty. Przy centromerach inny czynnik, Sgo1, rekrutuje fosfatazę, która usuwa tę grupę fosforanową, zachowując wiązanie Sororin–Pds5 i silne zablokowanie bramki aż do ostatecznego przecięcia RAD21 w anafazie. Testy genetyczne wykazały, że gdy Wapl zostaje usunięty, defekty spójności wynikające z zaburzenia interakcji CTR–bramka zanikają, podkreślając, że zasadniczą rolą Sororin jest osłanianie bramki wyjścia przed otwarciem napędzanym przez Wapl.
Co to oznacza dla stabilności genomu i chorób
Wspólnie te odkrycia przedstawiają Sororin jako strażnika o podwójnej funkcji: konkuruje z Wapl o dostęp do Pds5 i jednocześnie bezpośrednio zaciska bramkę wyjścia DNA kohezyny przez swój drobny CTR. Ta podwójna kontrola zapewnia, że spójność jest silna tam, gdzie trzeba — zwłaszcza przy centromerach — a jednocześnie może być stopniowo usuwana z ramion chromosomów we wczesnej mitozie, by umożliwić prawidłowe upakowanie i rozplątywanie. Ponieważ mutacje w kohezynie i jej regulatorach są częste w zaburzeniach rozwojowych i nowotworach, precyzyjnie zmapowana powierzchnia kontaktu między Sororinem, RAD21 i SMC3 uwypukla nowe miejsce podatności, gdzie drobne zmiany mogą destabilizować chromosomy — oraz potencjalny cel dla przyszłych terapii, które chciałyby modulować, jak ściśle utrzymywane jest nasze materiał genetyczny.
Cytowanie: Chen, Q., Yuan, X., Shi, M. et al. Sororin locks the DNA-exit gate of cohesin to preserve sister-chromatid cohesion. Nat Commun 17, 2284 (2026). https://doi.org/10.1038/s41467-026-70484-2
Słowa kluczowe: spójność chromatyd siostrzanych, kompleks kohezyny, Sororin, separacja chromosomów, stabilność genomu