Clear Sky Science · pl
Decyzje wzrokowo‑motoryczne przez konwergencję wielofunkcyjnych cech w tylnej części mózgu larwalnej zebrafinki
Jak maleńkie rybki wyjaśniają codzienne wybory
W każdej chwili nasze mózgi przetwarzają wiele informacji wzrokowych: dokąd coś się przesuwa, gdzie jest jasno lub ciemno i jak te wzory się zmieniają. To badanie wykorzystuje maleńkie larwy zebrafinki, aby odpowiedzieć na wielkie pytanie istotne także dla ludzi: gdy różne wskazówki wzrokowe sugerują sprzeczne kierunki, czy mózg wybiera zwycięzcę, czy też cicho sumuje wszystkie sygnały, by zdecydować o ruchu?
Obserwowanie, jak rybki decydują, w którą stronę popłynąć
Larwy zebrafinki są idealne do tego problemu, ponieważ są przezroczyste i można obrazować ich cały mózg podczas odbierania bodźców i ruchu. Autorzy zaprojektowali proste, lecz potężne ustawienie: pojedyncza rybka pływa swobodnie w okrągłym naczyniu, podczas gdy projektor pod spodem wyświetla dwa rodzaje wzorów wizualnych. Jeden to pole poruszających się kropek, które zazwyczaj skłaniają zwierzę do płynięcia z prądem — stabilizacyjna reakcja zwana odpowiedzią optomotoryczną. Drugi to różnica lewo‑prawo w natężeniu światła — jedna połowa świata wizualnego jaśniejsza od drugiej — co przyciąga zwierzę ku jaśniejszej stronie, zachowanie zwane fototaksją. Poprzez ostrożne łączenie tych wzorów, czasami zgodnych, czasami sprzecznych, zespół mógł zmierzyć, jak często każda rybka wybiera skręt w lewo lub prawo oraz jak szybko zapadają te decyzje.
Sumowanie sygnałów zamiast wybierania jednego zwycięzcy
Naukowcy porównali zachowanie z dwiema prostymi zasadami decyzyjnymi. W strategii „zwycięzca bierze wszystko” najsilniejszy sygnał — ruch lub światło — powinien całkowicie dominować, szczególnie gdy jest wyraźny i wiarygodny. W strategii „addytywnej” ruch i światło każdy w niewielkim stopniu popychają rybkę w kierunku, a faktyczny wybór odzwierciedla sumę tych impulsów. W badanej grupie ryb wzorce wyborów odpowiadały regule addytywnej: zmiana natężenia światła po jednej stronie przesuwała całą krzywą skrętów wywołanych ruchem w górę lub w dół, tak jakby odrębny efekt świetlny był po prostu dodawany. Gdy ruch i światło wskazywały ten sam kierunek, rybki były bardziej trafne i reagowały szybciej; gdy wskazywały przeciwne kierunki, wybory oscylowały wokół losowości, a czasy reakcji się wydłużały — zgodnie z sytuacją, w której dwie siły ciągną w przeciwne strony, zamiast bycia zdominowanym przez jedną z nich.
Trzy drogi wzrokowe kształtujące jedną decyzję
Patrząc dokładniej w czasie, zespół odkrył, że „światło” nie było jednym jedynym wpływem. Zachowanie ujawniło trzy oddzielne wkłady, które razem kształtowały każdy epizod pływania. Po pierwsze, sygnały ruchu były powoli integrowane: im dłużej kropki dryfowały w jednym kierunku, tym większe prawdopodobieństwo, że zwierzę skręci w tę stronę. Po drugie, stałe różnice w poziomie światła między stronami delikatnie przyciągały rybę ku jaśniejszej połowie. Po trzecie, nagłe zmiany światła — gdy jedna strona gwałtownie stawała się jaśniejsza lub ciemniejsza — krótkotrwale odpychały rybę od zmieniającej się strony, działając jak krótkotrwały bodziec odpychający. Zwięzły model matematyczny z tymi trzema składnikami, z których każdy miał własną siłę i skalę czasową, dokładnie przewidywał, jak decyzje o skręcie rozwijały się w czasie dla dziesiątek różnych kombinacji bodźców, nawet tych, które nie były użyte do dopasowania modelu.
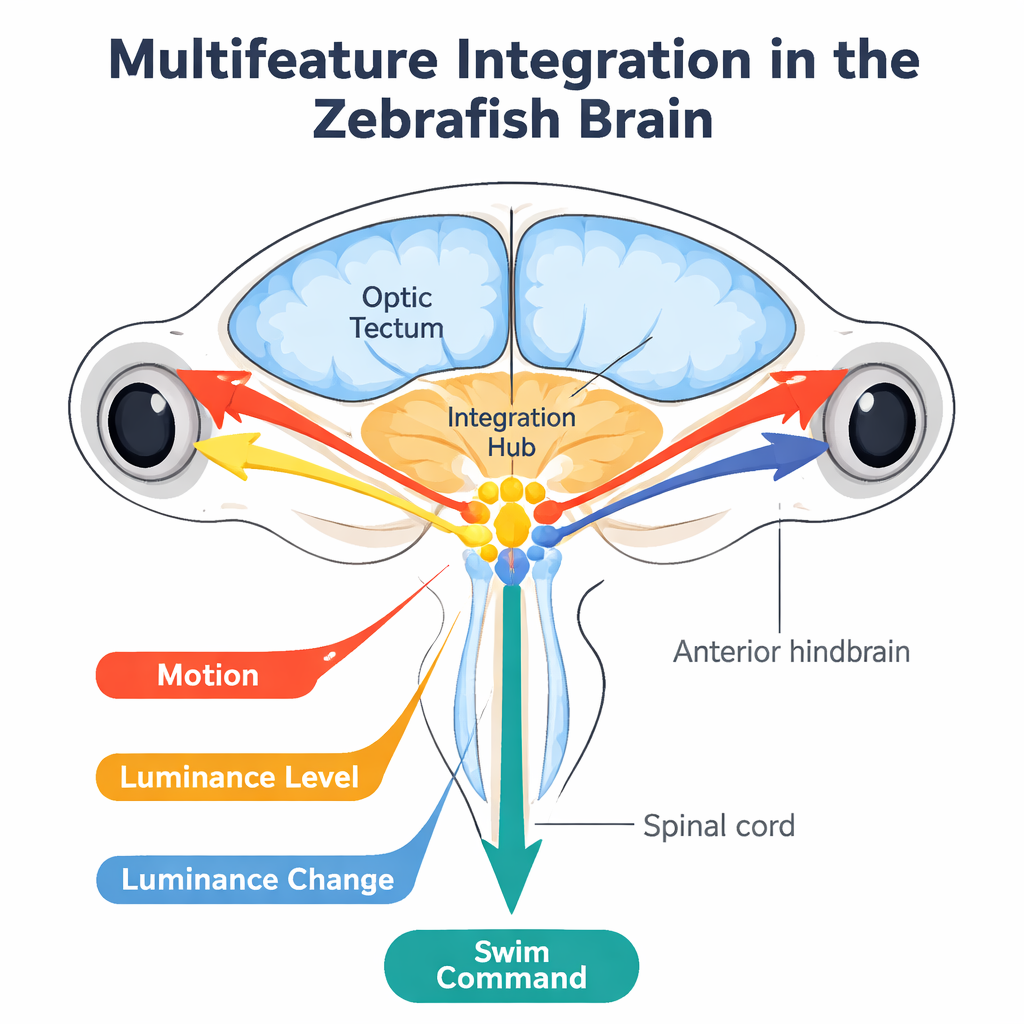
Znajdowanie ośrodka mózgowego, który łączy ruch i światło
Aby ustalić, gdzie zachodzą te obliczenia, autorzy użyli obrazowania wapniowego dwufotonowego obejmującego cały mózg, które rejestruje aktywność niemal wszystkich neuronów w żywej rybce. Prezentowali te same wzory ruchu i oświetlenia podczas nagrań i wyszukiwali komórki, których aktywność odpowiadała sygnałom przewidywanym przez model. Neurony reagujące na poziom światła i jego zmiany pojawiały się głównie w skrzyżowaniu wzrokowym (tectum opticum), ośrodku wzrokowym śródmózgowia, i w pokrewnych regionach. Komórki integrujące ruch oraz neurony, których aktywność odzwierciedlała ostateczny złożony „wielofunkcyjny” sygnał, skupiały się w części tyłomózgowia tuż za móżdżkiem. Dalsze eksperymenty, które znakowały neurony pobudzające i hamujące oraz śledziły kształty i projekcje pojedynczych komórek, wykazały w dużej mierze zrównoważone lokalne okablowanie z wieloma ścieżkami od oczu do tego przedniego tylnego ośrodka „integracyjnego” oraz wyjściami w kierunku obwodów motorycznych kontrolujących pływanie.
Od mózgów ryb do ogólnych zasad podejmowania decyzji
W codziennym życiu zwierzęta rzadko otrzymują jeden, perfekcyjnie wiarygodny sygnał. To badanie pokazuje, że przynajmniej w przypadku podstawowego przewodzenia wizualnego u zebrafinki mózg rozwiązuje ten problem, utrzymując ruch, jasność i zmianę jasności w częściowo oddzielnych kanałach, a następnie sumując je w dedykowanym ośrodku tylnego mózgu, by wygenerować decyzję o ruchu. Zamiast pozwalać jednemu sygnałowi wetować wszystkie pozostałe, układ działa jak prosty kalkulator, ważając każdą cechę zgodnie z jej siłą i czasem. Ponieważ podobne strategie addytywne pojawiają się u ssaków, w tym u ludzi, wyniki te sugerują, że wspólna, ogólnomózgowa zasada może leżeć u podstaw tego, jak różne kręgowce łączą sprzeczne informacje sensoryczne w spójne działania.
Cytowanie: Slangewal, K., Aimon, S., Capelle, M.Q. et al. Visuomotor decision-making through multifeature convergence in the larval zebrafish hindbrain. Nat Commun 17, 2024 (2026). https://doi.org/10.1038/s41467-026-69633-4
Słowa kluczowe: integracja multisensoryczna, zebrafinka, ruch wzrokowy, fototaksja, decyzje sensorimotoryczne