Clear Sky Science · pl
Dynamiczny sygnał kompromisu aktywność‑stabilność w ewolucji laktamazy
Dlaczego ma to znaczenie dla oporności na antybiotyki
Oporność na antybiotyki często wydaje się czarną skrzynką: bakterie „jakoś” się zmieniają i leki przestają działać. To badanie otwiera tę skrzynkę dla jednego z najlepiej poznanych enzymów oporności, TEM‑1 β‑laktamazy, który pozwala bakteriom niszczyć antybiotyki podobne do penicyliny. Śledząc, jak kształt i ruchy tego enzymu zmieniają się w miarę ewolucji pod kątem nowszych leków, autorzy wyjaśniają, dlaczego mutacje zwiększające aktywność często osłabiają stabilność i jak ewolucja znajduje sprytne sposoby zrównoważenia obu cech.
Od niszczyciela penicyliny do szerszego łamacza leków
TEM‑1 pierwotnie świetnie radzi sobie z rozkładem starszych antybiotyków penicylinowych, ale słabiej z nowszymi, bardziej masywnymi lekami, takimi jak cefotaksym. W wielu szczepach klinicznych pojawia się pojedyncza kluczowa mutacja nazwana G238S w pobliżu miejsca aktywnego enzymu, kieszeni, w której antybiotyki są cięte. Ta mutacja znacznie poprawia zdolność enzymu do niszczenia cefotaksymu, przy jednoczesnym umiarkowanym pogorszeniu jego pierwotnej aktywności wobec penicylin. Autorzy pokazują, że G238S nie tylko rozszerza kieszeń; reorganizuje sposób poruszania się kilku otaczających pętli i helis, tworząc nową funkcjonalną konformację lepiej dopasowaną do masywnych cząsteczek leków.
Ruch białka jako ewolucyjny pokrętło strojenia
Wykorzystując zaawansowane techniki rezonansu magnetycznego jądrowego (NMR), badacze zmierzyli, jak różne części TEM‑1 poruszają się w skalach czasowych od trylionowych do tysięcznych części sekundy. Dzika forma TEM‑1 jest dość sztywna, co pomaga jej wydajnie przetwarzać pierwotne substraty. G238S pozostawia większość tej szybkiej sztywności nienaruszoną, ale wprowadza wolniejsze, precyzyjnie dostrojone ruchy w wielu ścianach miejsca aktywnego. Ruchy te są na tyle szybkie, by nadążyć za chemią enzymatyczną, a jednocześnie niezbyt gwałtowne, by zaburzyć krytyczne reszty katalityczne. Efektem jest „optymalne okno” elastyczności: wystarczająco ruchu, by otworzyć kieszeń dla cefotaksymu, przy jednoczesnym utrzymaniu właściwego ustawienia głównego aparatu chemicznego.
Zrównoważenie dwóch kształtów zamiast wybierania jednego
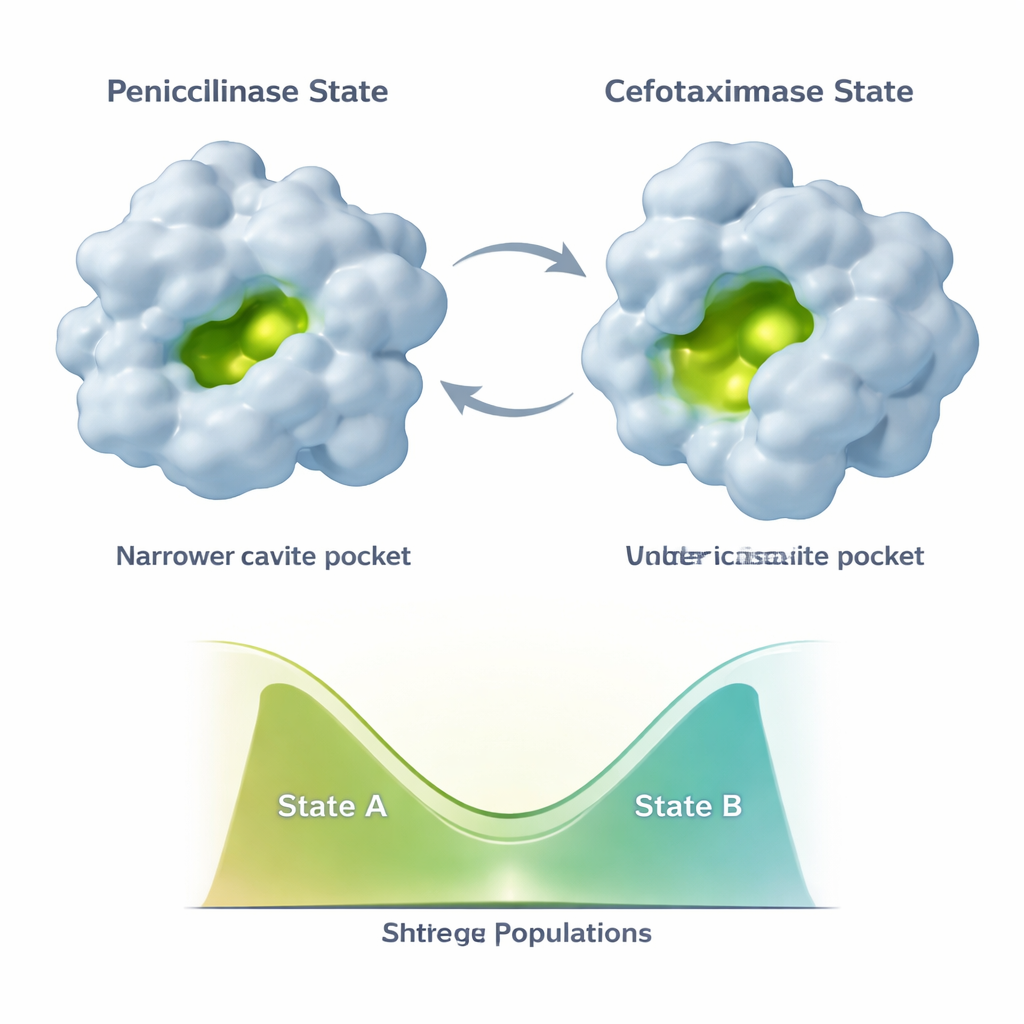
Ewolucja nie zamraża TEM‑1 w jednej nowej formie. Zamiast tego enzym próbuje przynajmniej dwóch głównych konformacji: stanu „penicylinazy” przypominającego strukturę pierwotną oraz bardziej otwartego stanu „cefotaksymazy” lepiej przystosowanego do nowszych leków. Dodatkowe mutacje pojawiające się później, takie jak E104K i A42G, robią coś subtelnego. Zamiast po prostu stabilizować nową, przyjazną cefotaksymowi formę, przesuwają proporcje między tymi dwoma stanami. Dane NMR pokazują, że różne części miejsca aktywnego i wspierającego rusztowania mogą niezależnie przesuwać swoje populacje wzdłuż tego kontinuum dwóch stanów. Powstaje w ten sposób kombinatoryczny zestaw wariantów enzymu, z których każdy ma inną mieszankę konformacji przypominających penicylinę i cefotaksym, a co za tym idzie różne profile katalityczne.
Ukryte słabe punkty i naprawy na odległość
Mutacje poprawiające aktywność często niosą ukryty koszt: czynią białko mniej stabilnym. Zamiast patrzeć tylko na całkowite rozwinięcie, zespół odwzorował lokalną stabilność na poziomie krótkich segmentów przy użyciu wymiany wodór–deuter z detekcją spektrometrii mas. Stwierdzono, że G238S destabilizuje nie tylko pobliskie pętle, ale też odległe helisy i regiony β‑kartki tworzące szkielet strukturalny. Niektóre z tych regionów nachodzą na „kryptyczną” kieszeń allosteryczną — rzadko odwiedzane otwarcie w rdzeniu białka, które może wiązać małe cząsteczki i tłumić aktywność. G238S ułatwia otwarcie tej kieszeni, skutecznie wbudowując łagodną cechę samoinhibicyjną w enzym. Późniejsze mutacje, szczególnie A42G, wzmacniają tę osłabioną sieć szkieletu, poprawiając lokalną stabilność wokół trzech oddziałujących helis bez usuwania korzystnej dynamiki miejsca aktywnego. Innymi słowy, ewolucja łatwiejszych naprawia odległe słabe punkty struktury, zamiast cofać pierwotnej innowacji.
Co badanie ujawnia o strategii ewolucji
Dla czytelnika nietechnicznego główne przesłanie jest takie, że białka takie jak TEM‑1 nie rozwijają oporności przez proste przełączanie włącz/wyłącz. Każda mutacja nieznacznie zmienia sposób, w jaki enzym „oddycha”, wygina się i dzieli czas między różne aktywne pozy. G238S otwiera drzwi do nowego zadania — rozkładu nowszych antybiotyków — ale jednocześnie tworzy lokalne kruche miejsca i częściowo samoinhibujący stan. Mutacje wtórne działają jak przemyślane wzmocnienia, stabilizując rusztowanie i dopracowując równowagę między starymi i nowymi konformacjami, dzięki czemu enzym może pozostać zarówno aktywny, jak i trwały. Takie dynamiczne spojrzenie na ewolucję, w którym ruchy i lokalne słabości mają taką samą wagę jak struktura statyczna, może pomóc w projektowaniu przyszłych antybiotyków i leków celujących w enzymy, na które bakterie trudniej będzie znaleźć odpowiedź.
Cytowanie: Arcia, E., Keramisanou, D., Jacobs, L.M.C. et al. Dynamic signature of activity-stability tradeoff in lactamase evolution. Nat Commun 17, 1884 (2026). https://doi.org/10.1038/s41467-026-68620-z
Słowa kluczowe: oporność na antybiotyki, beta‑laktamaza, ewolucja białek, dynamika enzymu, stabilność białka