Clear Sky Science · pl
Różne role podtypów komórek warstwy 5 kory w uczeniu asocjacyjnym
Jak mózg uczy się, że dotyk zapowiada nagrodę
Wyobraź sobie, że uczysz się, iż delikatne stuknięcie w nadgarstek oznacza, że będzie deser. Twój mózg musi powiązać prosty dotyk z przyszłą nagrodą. Badanie to zagląda do mózgu myszy, aby zobaczyć, jak dwa różne typy komórek nerwowych współdziałają przy tworzeniu takich powiązań, ujawniając, jak codzienne uczenie się dzieli się na „co się stało” i „co to dla mnie znaczy”. 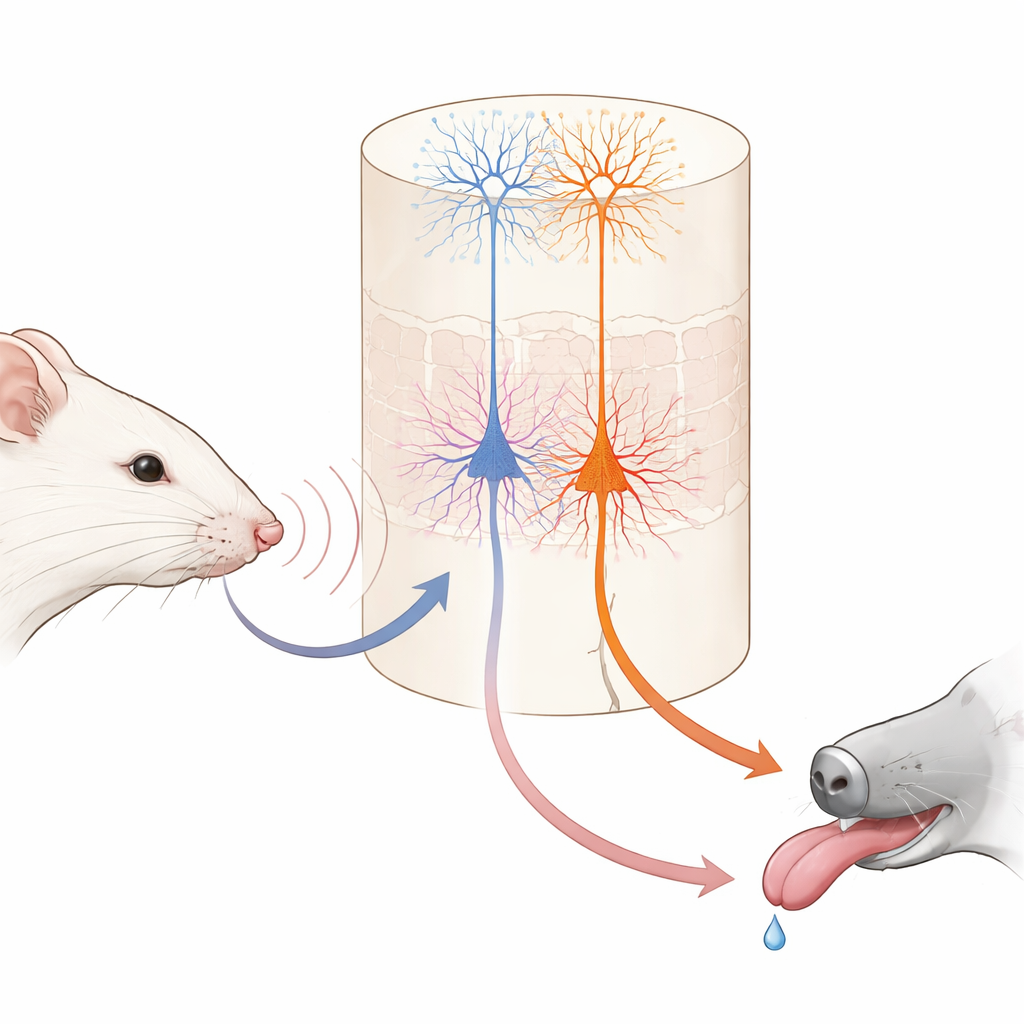
Prosta gra w wąsy służąca do badania uczenia się
Naukowcy trenowali myszy z unieruchomioną głową w prostą grę wykorzystującą wibrujące wąsy twarzy. W każdym podejściu pojedynczy wąs był wprawiany w drganie jedną z dwóch prędkości. Jedno drganie było po krótkim opóźnieniu nagradzane maleńką kroplą wody; drugie nigdy nie dawało nagrody. Na początku myszy liżały w oczekiwaniu przy obu rodzajach prób, nie wiedząc jeszcze, które odczucie na wąsie sygnalizuje wodę. W ciągu kilku dni stopniowo nauczyły się wczesnego lizania tylko przy „dobrej” wibracji i powstrzymywania się przy tej nienagradzanej. Gdy naukowcy tymczasowo wyciszali podstawowy obszar dotykowy kory podczas treningu, to uczenie w dużej mierze znikało, co pokazuje, że ten obszar sensoryczny jest ważny dla budowania skojarzenia, choć doświadczone myszy później mogły wykonywać zadanie bez niego.
Dwa typy komórek wyjściowych o bardzo różnych zadaniach
W obrębie tego obszaru dotykowego głęboka warstwa znana jako warstwa 5 zawiera dwa główne rodzaje komórek wyjściowych. Jedna grupa, tu nazwana komórkami IT, wysyła sygnały do innych obszarów korowych w obu półkulach mózgu. Druga grupa, komórki ET, przekazuje sygnały głównie w dół do struktur podkorowych zaangażowanych w działanie i nagrodę. Dzięki genetycznie zmodyfikowanym myszom i wysokorozdzielczej mikroskopii dwu-fotonowej autorzy mogli selektywnie monitorować aktywność każdego typu komórek przez końcówki ich długich, drzewkowatych gałęzi. Przed nauką komórki IT już silnie i niezawodnie reagowały na wibracje wąsów, a ich łączna aktywność mogła dokładnie rozróżnić dwie prędkości drgań. Komórki ET w przeciwieństwie do nich reagowały słabiej i mniej konsekwentnie na bodziec, dając jedynie rozmyty odczyt, które drganie miało miejsce.
Stabilne wrażenia kontra rosnące oczekiwania
W miarę jak myszy się uczyły, komórki IT zachowywały się jak wiarygodni raporterzy. Ich odpowiedzi pozostawały ściśle związane z momentem ruchu wąsa i zmieniały się niewiele z dnia na dzień. Nadal kodowały, które drganie wystąpiło, niezależnie od tego, czy przewidywało ono nagrodę. Komórki ET natomiast przekształciły swoje działanie. Zamiast po prostu iskrować przy początku bodźca, ich aktywność stopniowo narastała podczas wibracji i krótkiego opóźnienia, osiągając szczyt wokół oczekiwanego momentu podania wody. To narastanie rosło równolegle z lizaniem w oczekiwaniu u zwierząt i stawało się lepsze w przewidywaniu, czy próba zakończy się lizaniem, niż w raportowaniu dokładnego bodźca. Pojedyncze komórki ET wchodziły i wychodziły z aktywnej grupy w ciągu dni, ale na poziomie populacji wzorzec stawał się bardziej spójny, co sugeruje elastyczny, lecz zbieżny kod oczekiwania nagrody. 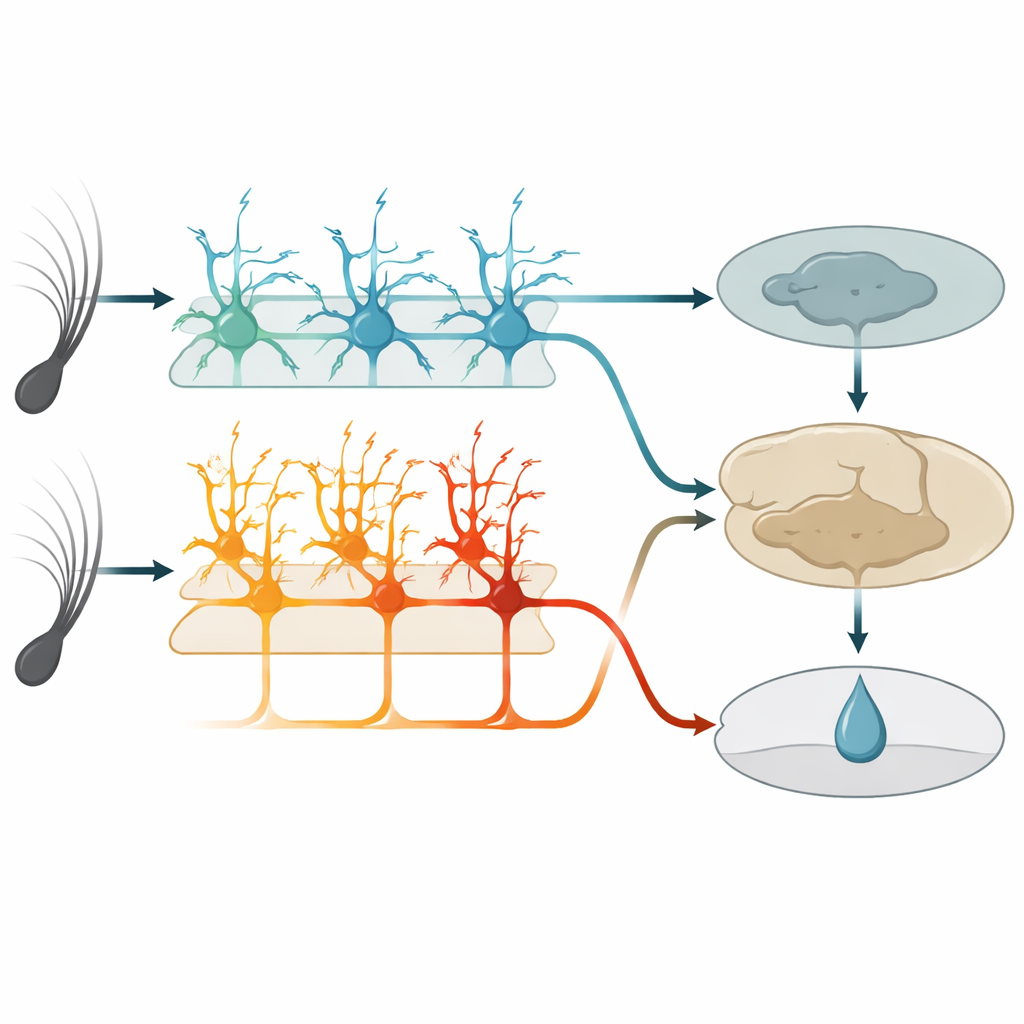
Wyłączanie każdego typu komórek ujawnia podział pracy
Aby przetestować funkcję, zespół użył narzędzi chemogenetycznych, by selektywnie osłabić albo komórki IT, albo ET podczas treningu. Gdy komórki IT były wyciszone, myszy ogólnie wykazywały mniej lizania w oczekiwaniu i nie potrafiły zbudować wyraźnej różnicy między drganiami nagradzanymi i nienagradzanymi. Gdy wyciszono komórki ET, myszy zachowywały się odwrotnie: zbytnio lizły przy obu drganiach, zwłaszcza przy tym nienagradzanym, i miały trudności z dopracowaniem zachowania, choć nadal energicznie liżały. Wyciszenie którejkolwiek grupy po opanowaniu zadania nie szkodziło już wykonaniu, co sugeruje, że gdy inne obszary mózgu zapiszą skojarzenie, ten obszar sensoryczny i jego wyjścia z warstwy 5 stają się mniej krytyczne dla realizacji wyuczonej reakcji.
Model uczenia się odzwierciedlający zachowanie mózgu
Autorzy zbudowali model komputerowy w stylu uczenia przez wzmocnienie, aby zinterpretować te odkrycia. W modelu sieć podobna do IT dostarcza stabilnych reprezentacji sensorycznych, które pomagają oszacować „wartość” każdego bodźca — jak prawdopodobne jest, że zostanie on nagrodzony. Droga podobna do ET przekazuje tę przewidywaną wartość do obwodu niższego rzędu, który porównuje ją z rzeczywistą nagrodą, generując błąd predykcji, który koryguje przyszłe estymaty wartości. Zablokowanie ścieżek IT lub ET w modelu odtworzyło odrębne deficyty uczenia obserwowane u myszy: bez wejścia typu IT uczenie było powolne i słabe dla obu bodźców; bez wyjścia typu ET początkowe uczenie się następowało, ale system nie potrafił odpowiednio zmniejszyć reakcji na bodziec nienagradzany. Model oddał też, jak z czasem obszary nie-sensoryczne mogą przejąć wykonanie zadania, zgodnie z eksperymentami.
Co to oznacza dla codziennego uczenia się
Mówiąc prosto, badanie sugeruje, że gdy uczymy się, iż konkretny obraz, dźwięk lub dotyk zapowiada coś dobrego lub złego, różne zestawy głębokich neuronów kory dzielą między siebie pracę. Jedna grupa zachowuje wierny zapis „co się stało” w świecie, podczas gdy druga stopniowo zaczyna sygnalizować „czego spodziewam się następnym” i pomaga dopracować zachowanie poprzez porównanie tych oczekiwań z rzeczywistością. Razem tworzą most między surowym doznaniem a elastycznymi, opartymi na doświadczeniu działaniami, oferując jaśniejszy obraz tego, jak mózg wspiera nawyki, uczenie się z nagród i — być może — jak te procesy ulegają zaburzeniom w chorobie.
Cytowanie: Moberg, S., Garibbo, M., Mazo, C. et al. Distinct roles of cortical layer 5 subtypes in associative learning. Nat Commun 17, 2648 (2026). https://doi.org/10.1038/s41467-026-68307-5
Słowa kluczowe: uczenie asocjacyjne, kora somatosensoryczna, neurony warstwy 5, oczekiwanie nagrody, uczenie przez wzmocnienie