Clear Sky Science · nl
Moleculaire basis van antagonisme van de dimerische humane arginine-vasopressine receptor 1A
Waarom een hormoonreceptor in de hersenen ertoe doet
Hormonen zoals vasopressine en oxytocine zijn vooral bekend om hun rol in het regelen van waterbalans, bloeddruk, bevalling en hechting. Maar hoe hun receptoren op atomair niveau functioneren bleef grotendeels verborgen. Dit artikel onthult gedetailleerde driedimensionale structuren van één belangrijke receptor, de humane vasopressine V1a-receptor, die gekoppeld is aan sociaal gedrag, stress en verschillende hersenaandoeningen. Inzicht in de vorm van deze receptor en in hoe geneesmiddelen deze blokkeren kan onderzoekers helpen betere behandelingen te ontwerpen voor aandoeningen zoals autisme, posttraumatische stressstoornis en de ziekte van Huntington.
Een tweelingreceptor die hart-, nier- en hersensignalen vormt
De V1a-receptor bevindt zich op het oppervlak van veel cellen in het lichaam, vooral in bloedvaten, nieren en bepaalde hersengebieden. Wanneer het hormoon vasopressine eraan bindt, activeert de receptor interne signaalroutes die de bloeddruk, de vochtbalans en hersencircuits voor sociale interactie, emotie en stress reguleren. Genetische en klinische studies hebben afwijkende V1a-signaaloverdracht in verband gebracht met autismespectrumstoornissen, PTSS en de ziekte van Huntington, waardoor het een aantrekkelijk doelwit voor geneesmiddelen is. Verschillende V1a-blokkers (antagonisten) zijn al in gebruik of in klinische proeven, maar tot nu toe had niemand de humane V1a-receptor met hoge resolutie gezien, waardoor belangrijke vragen over hoe hij assembleert en hoe precies deze geneesmiddelen hem uitschakelen onbeantwoord bleven. 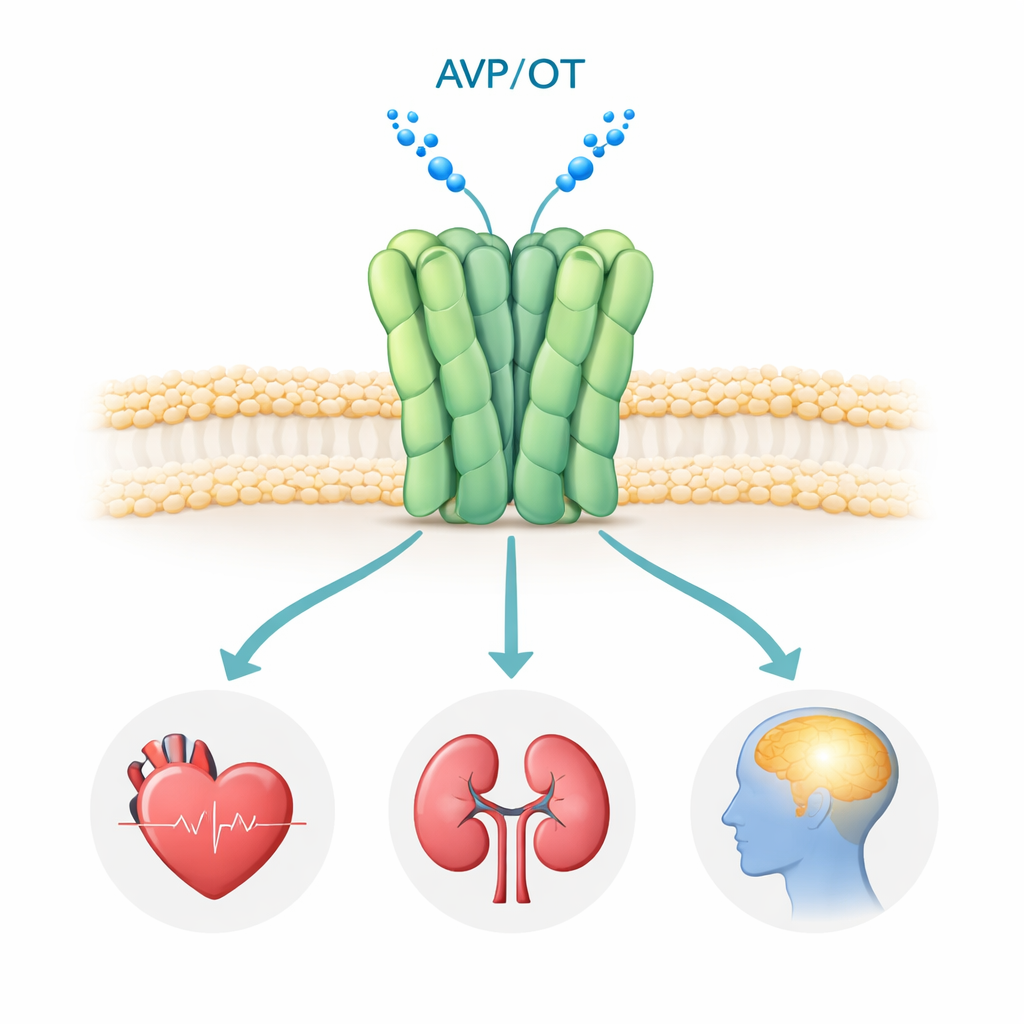
De structuur van de receptor vastleggen in meerdere geneesmiddelgebonden toestanden
De onderzoekers gebruikten cryo-elektronenmicroscopie, een techniek waarbij eiwitten vlug worden ingevroren en met een elektronenbundel worden afgebeeld, om de humane V1a-receptor te visualiseren. Om het eiwit te stabiliseren maakten ze een licht aangepaste vorm die nog goed aan geneesmiddelen bindt en combineerden die met een klein antistoffragment (een nanodeeltje) om de beeldvorming te vergemakkelijken. Ze bepaalden structuren van de receptor alleen en gebonden aan drie medisch belangrijke antagonisten: atosibaan (een peptidemiddel dat wordt gebruikt om vroegtijdige bevalling te voorkomen) en twee brein-penetrerende kleine moleculen, balovaptan en SRX246, die in mensen met autisme of de ziekte van Huntington zijn getest. Alle structuren bereikten bijna-atomaire resolutie en toonden de posities van de zeven membraan-doorstekende helices van de receptor, flexibele lussen en de gebonden geneesmiddelen.
Een receptor die bij voorkeur in paren werkt
In tegenstelling tot verwante receptoren die alleen als afzonderlijke eenheden waren gezien, verscheen V1a als een paar — een dimer — in alle vier de cryo-EM-structuren. De twee receptoren liggen naast elkaar in het membraan en maken strakke contacten voornamelijk via één van hun helices, geholpen door zowel polaire als vettige (hydrofoob) interacties. Om te testen of dit koppelen ook in levende cellen voorkomt, fuseerde het team V1a met een helder fluorescerend eiwit en gebruikte een enkel-molecuul fotoblekingstechniek: als een plek op het celoppervlak twee receptorkopieën bevatte, zou het licht in twee stappen verdwijnen. Ongeveer driekwart van de waargenomen vlekken verbleekte in precies twee stappen, wat sterk ondersteunt dat V1a van nature dimers vormt aan het celoppervlak. Wanneer de wetenschappers sleutelresiduen die het contact vormen muteerden om het interface te verstoren, verschoof de receptor naar enkele eenheden en werd hij minder reactief op hormoon en geneesmiddel, wat suggereert dat de dimer niet slechts structurele versiering is maar functioneel belangrijk.
Een flexibele poort bij de hormoeningang
Het team ontdekte een onverwacht “hek”-gebied dat extracellulaire lus 2 (ECL2) wordt genoemd en zich bovenin de hormoonbindingspocket bevindt. In de geneesmiddelvrije (apo-)toestand ligt deze lus plat over de pocket als een deksel en vormt hij niet de gebruikelijke disulfidebinding (een zwavel–zwavelverbinding) die bij veel verwante receptoren wordt gezien. In plaats daarvan vouwen delen van de lus zich in de pocket en worden ze op hun plaats gehouden door een web van interacties met omliggende helices, waardoor ze de grote, kleverige bindingsholte gedeeltelijk afdekken. Wanneer een van de drie antagonisten bindt, zwaait ECL2 omhoog en weg, vormt de klassieke disulfidebinding en ontstaat er een brede, door oplosmiddel gevulde holte die de geneesmiddelen innemen. Deze dramatische beweging suggereert dat V1a ECL2 mogelijk gebruikt als een dynamische barrière om willekeurige activatie door toevallige moleculen te beperken, en dat geneesmiddelen kunnen worden ontworpen om de lus ofwel in zijn platte “grondtoestand” te vergrendelen ofwel zijn opgeheven, open conformatie te benutten.
Hoe drie geneesmiddelen dezelfde receptor op verschillende manieren stilleggen
Atosibaan, dat sterk het natuurlijke hormoon oxytocine nabootst, reikt van de top van de pocket tot de basis en verankert zich via een combinatie van waterstofbruggen en hydrofobe contacten. Door enkele sleutelposities ten opzichte van oxytocine te veranderen, slaagt het er niet in de keten van interne verschuivingen teweeg te brengen die normaal nodig zijn voor receptoractivatie: cruciale “microswitch”-residuen die tijdens signalering bewegen blijven vergrendeld in hun inactieve posities, de interne holte die het G-eiwit toeliet opent nooit, en een magnesium-bindingsplaats die belangrijk is voor activatie wordt verstoord. Daarentegen zijn balovaptan en SRX246 compacte, niet-peptide moleculen die diep in de pocket dringen maar verschillende strategieën gebruiken. Balovaptan vertrouwt op een rigide, hydrofoob kern die strak in een diepe spleet past, plus een flexibele polaire staart die naar de pocketingang reikt. SRX246 gebruikt een modulaire, fragmentachtige architectuur verankerd door een β-lactamkern, met verschillende “zones” die subpockets opvullen en zich uitstrekken richting de extracellulaire lussen. In beide gevallen stabiliseren de geneesmiddelen een inactieve conformatie die onverenigbaar is met binding van het G-eiwit. Subtiele verschillen in pocketvorm en chemie — vooral op twee posities op helices 5 en 7 — helpen verklaren waarom balovaptan en SRX246 V1a prefereren boven nauw verwante receptoren.
Gevolgen voor toekomstige therapieën
Door hoge-resolutie momentopnamen van V1a als dimer te leveren, een eerder ongeziene “platte” lusconformatie in de geneesmiddelvrije toestand te onthullen en te laten zien hoe drie zeer verschillende antagonisten de receptor uitschakelen, biedt dit werk medicijnontwerpers een precies structureel handboek voor het richten op V1a. Het suggereert manieren om volgende generaties geneesmiddelen te ontwerpen die ofwel dimer-specifieke kenmerken benutten of de receptor vergrendelen in een bijzonder inactieve grondtoestand, met als uiteindelijke doel hersen- en stressgerelateerde stoornissen specifieker te behandelen en met minder bijwerkingen.
Bronvermelding: Zhong, P., Chu, B., Yu, Z. et al. Molecular basis of antagonism of the dimeric human arginine vasopressin receptor 1A. Nat Commun 17, 1622 (2026). https://doi.org/10.1038/s41467-026-68331-5
Trefwoorden: vasopressine V1a-receptor, G-eiwitgekoppelde receptor, receptor-dimerisatie, cryo-EM-structuur, ontwerp van neuropsychiatrische geneesmiddelen