Clear Sky Science · ja
エンテロウイルスA群およびB群におけるディープミューテーショナルスキャンデータの比較解析は機能的分岐と治療標的を同定する
小さなRNAウイルスが私たちにとって重要な理由
エンテロウイルスは世界中に静かに循環する小さなRNAウイルスの大きな群で、通常は軽い風邪や胃腸炎を引き起こしますが、時に麻痺や心臓障害など重篤な病態を招くことがあります。これらに広く効くワクチンはほとんどなく、効果的な薬もさらに少ないのは、ウイルスの進化が非常に速いためです。本研究は一見単純だが実用的に重要な問いを投げかけます:ウイルスが変異するとき、どの部位は自由に変わりうるのか、どの部位は進化によってほとんど固定されているほど重要なのか?その答えはウイルスが容易に回避できない治療戦略の設計へとつながります。
ウイルスの設計図を一変異ずつ読み解く
研究者たちは非常に異なる疾患を引き起こす二つのヒトエンテロウイルスに焦点を当てました:小児の重篤な神経疾患に関連するエンテロウイルスA71と、心筋炎や膵臓がんに関連するコクサッキーウイルスB3です。ディープミューテーショナルスキャンという手法を用い、各ウイルスのタンパク質のほぼすべての位置を体系的に変えたウイルスライブラリを作成しました。これらの変異ウイルスを培養細胞に感染させ、高速シーケンシングで各変化がウイルス増殖にどう影響するかを測定しました。各変異体を元のウイルスと比較することで、ウイルスプロテオームのどの部位が変化を許容し、どの部位が強く制約されるかの詳細な地図を作成しました。 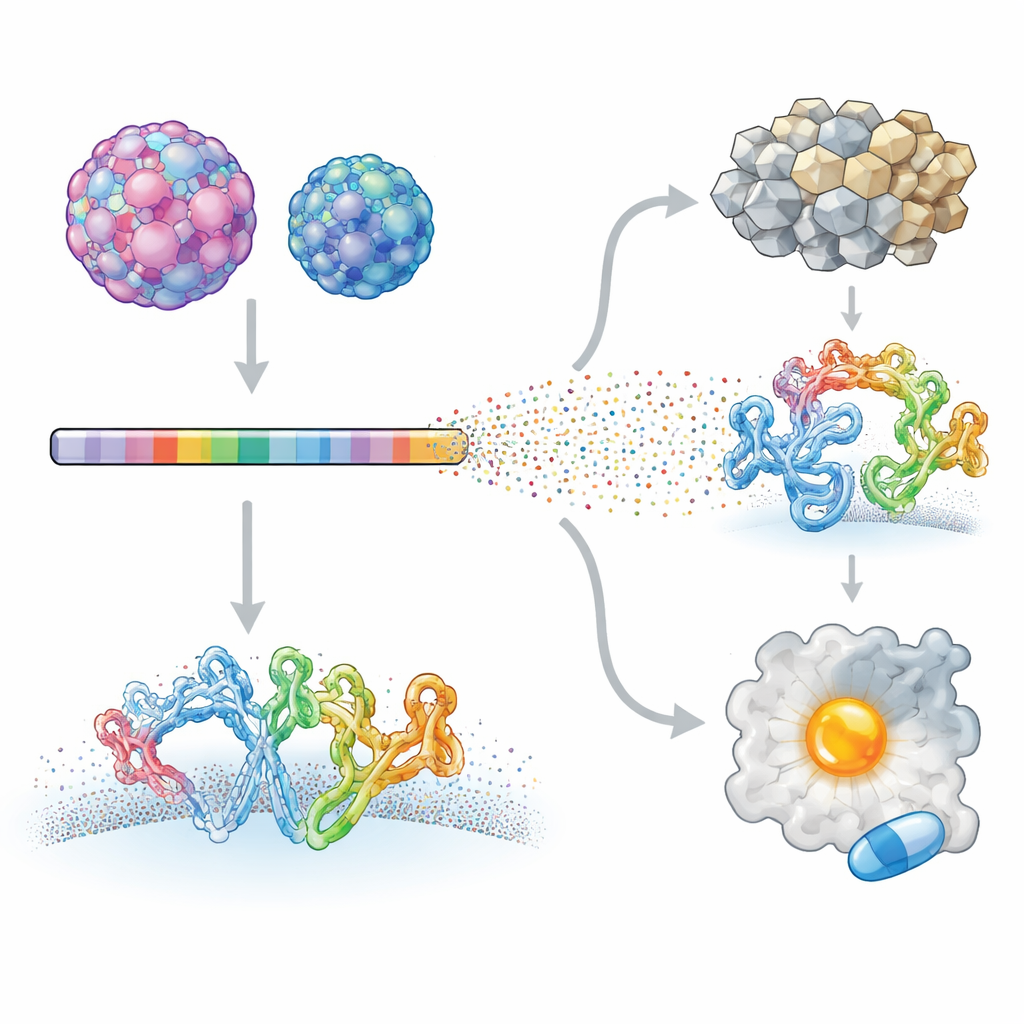
共通の強い制約とウイルス固有の柔軟性
アミノ酸配列が約半分しか一致しないにもかかわらず、両ウイルスは制約の全体像で著しく似たパターンを示しました。ゲノムを複製し、ウイルスタンパク質を切断し、RNAをほどく内部の酵素的 ‘‘働き手’’ は両方のウイルスで変異に非常に敏感でした。同様に、タンパク質殻(カプシド)を結びつける隠れた表面も、ウイルスを壊さずに変えることが難しいことがわかりました。対照的に、多くの外向きの領域や宿主と相互作用する領域はより変化を許容し、しばしば二種間で大きく異なっていました。構造解析は、変異に耐えられない保存された領域が触媒部位や組み立て界面の周辺に集積する一方、宿主受容体や膜、免疫分子に接する領域は種特異的な柔軟性のホットスポットであることを示しました。
ウイルスが細胞に接触し防御をかわす仕組み
研究チームは次にウイルスと宿主の分子レベルの“握手”に注目しました。エンテロウイルスA71とコクサッキーウイルスB3は侵入のために異なる細胞受容体を利用しており、研究ではカプシド表面の接触フットプリントが変異許容性の点で最も発散している領域の一つであることがわかりました。あるウイルスで受容体をつかむ残基はそのウイルスでは強く制約される一方で、もう一方のウイルスでは比較的許容的であり、それぞれがドッキングプラットフォームを微調整してきたことを反映しています。類似の分岐は、細胞膜を再構築して複製のための宿主因子を動員する小さな膜結合タンパク質3Aでも現れました。モデリングは、両ウイルスが同じ宿主因子GBF1に結合するために部分的に異なる接触パッチを3A上で使い、膜への挿入深さも異なる可能性を示唆しました。これらの違いは、近縁のウイルスが異なる組織を感染し、異なる病態を引き起こす理由を説明する助けになります。
実験室での進化が見落とすもの—そして正しく捉えるもの
培養細胞実験を現実世界の文脈に置くために、著者らは変異地図を患者試料由来の何千もの自然ウイルス配列と比較しました。全体として、実験室で柔軟とされた部位は自然界でも変動しやすく、特に種レベルの広い比較で顕著でした。しかし、各部位でどの特定のアミノ酸が好まれるかを詳しく調べると、自然進化と実験室選択は一つのウイルスタイプ内で最も密接に一致しました。注目すべきは、最大の不一致が外側カプシド表面や非構造タンパク質2Aおよび3Aの宿主相互作用領域に集中していた点で、まさに免疫応答や複雑な宿主環境が最も影響することが予想される場所でした。これはディープミューテーショナルスキャンがウイルスタンパク質の基本的な機械的制約を捉える一方で、実際の感染は免疫や組織嗜好性(トロピズム)からくる追加の圧力を上に重ね、これらはin vitroで再現しにくいことを示唆します。
将来の薬剤のための共通の弱点を見つける
最後に、これらの地図が薬剤開発の共通のアキレス腱を示せるかを検討しました。計算ツールを使って、二つのウイルスのタンパク質表面に小分子薬が結合しうるポケットを探索し、さらに変異データを重ね合わせてウイルスが自己を損なうことなく容易に変えられない残基から構成されるポケットを特定しました。その中で際立っていたのが2Cヘリカーゼ上のポケットでした。2CはウイルスRNAをほどくリング状のモーターで、その形状と構成残基は四つの主要なヒトエンテロウイルス種にわたって高度に保存されており、既知の活性部位から離れた位置にあり、自然界でも変化が稀で実験室テストでは変化が致命的であることが示されました。 
将来の治療への意味
二つの重要なエンテロウイルスについて可能な単一塩基変化がそれぞれどのように影響するかを体系的に調べることで、本研究はウイルスの共有され保存された硬直したコアと宿主とのより柔軟でウイルス特異的なインターフェースとの明確な分離を明らかにしました。保存されたコアには、変異によってウイルスの適合度を損なわせやすいことが示された2Cヘリカーゼ上の新たに注目されたポケットが含まれ、耐性障壁の高い広域抗ウイルス薬の有望な標的となります。同時に、より適応性の高い外側表面や宿主接触部位は、関連ウイルスが体内で非常に異なる振る舞いを示す理由を説明し、ワクチンや免疫ベースの治療が高速な進化に対処しなければならない領域を指し示します。これらの知見は、ウイルス自身の進化的限界を利用する治療設計のためのロードマップを提供します。
引用: Álvarez-Rodríguez, B., Bakhache, W., McCormick, L. et al. Comparative analysis of deep mutational scanning datasets in enteroviruses A and B identifies functional divergence and therapeutic targets. Nat Ecol Evol 10, 467–480 (2026). https://doi.org/10.1038/s41559-026-02993-8
キーワード: エンテロウイルス, ウイルス進化, ディープミューテーショナルスキャン, 抗ウイルス標的, 2Cヘリカーゼ