Clear Sky Science · ja
異種起源の細胞小器官を利用するための一時的分子キメラリズム
単一細胞が借りるソーラーパネル
太陽エネルギーを利用する生命と聞けば通常は植物や藻類を思い浮かべますが、単細胞の捕食者の中には近道をする者がいます:餌から「ソーラーパネル」――葉緑体――を盗むのです。本研究は、そのような微生物のひとつRapaza viridisが、異種由来のこれらの部品を数週間にわたってどのように稼働させ続けるかを探ります。宿主自身のタンパク質がどのようにして外来の葉緑体に入り込み、それらを支えるかを明らかにすることで、葉緑体のような複雑な細胞小器官が古くにどのようにして初めて進化したかについての洞察を与えます。
微生物はどうやって緑の力を盗んで使うか
Rapaza viridisは微小な鞭毛虫で、特定の緑藻Tetraselmisを摂食します。すべてを消化するのではなく、Rapazaは捕獲した葉緑体(ここでは「クレプトプラスト」と呼ばれる)を保持し、核を含む残りの藻類細胞は廃棄します。これらのクレプトプラストは細切れにされて娘個体へ受け継がれます。およそ2週間にわたり、宿主はこれら借用した葉緑体が生み出すエネルギーと炭素だけでほぼ生活できるほどで、元の藻類ゲノムからの指示や補修部品を受け取らなくても機能し続けます。この特異な生活様式は、恒久的な葉緑体への初期段階がどのようであったかを生きたまま覗く窓を提供します。
宿主遺伝子が盗まれた部品を維持する
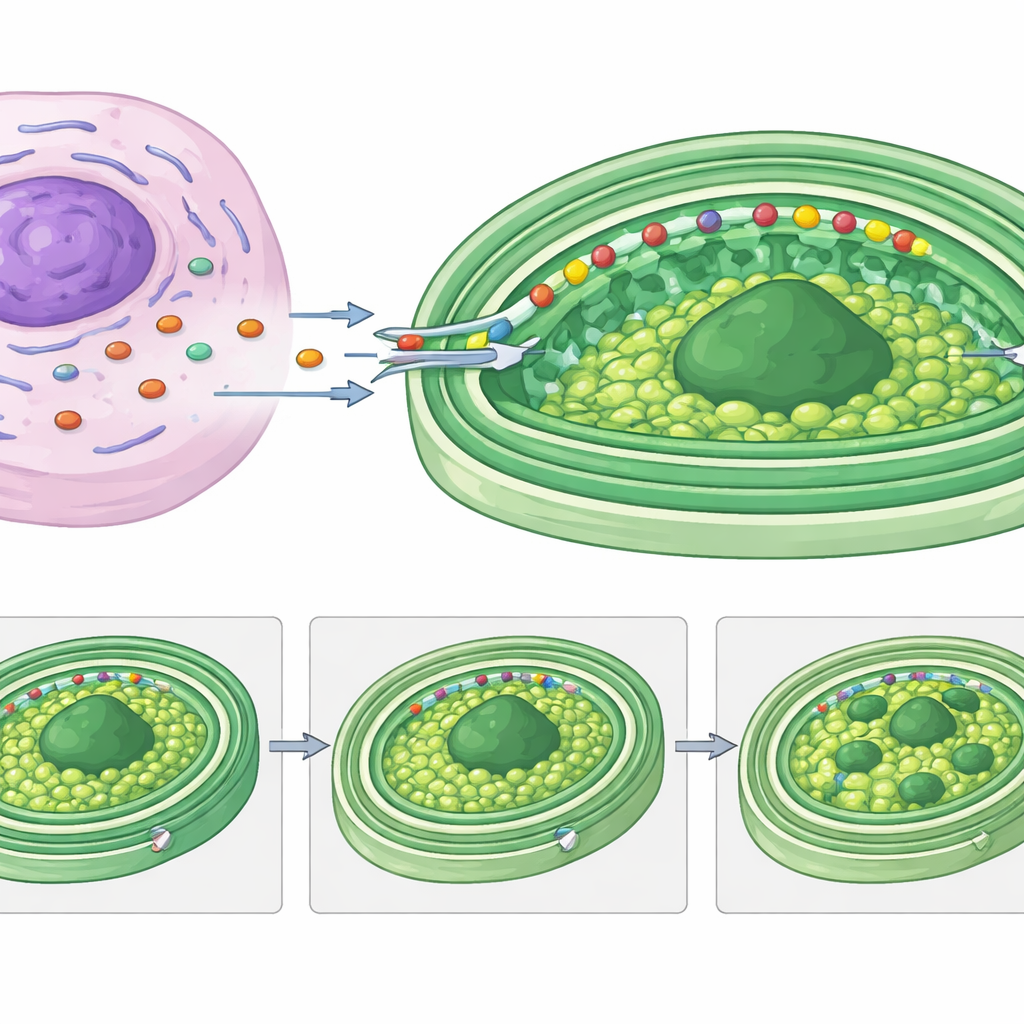
研究者たちは重要な疑問を投げかけました:藻類の核が失われたなら、クレプトプラストを稼働させるために必要なタンパク質は誰が供給するのか?Rapazaの遺伝子発現を時間軸で解析したところ、葉緑体内で働くと思われる宿主由来の遺伝子37個を同定しました。これらの多くは光捕集タンパク質、電子伝達成分、炭素固定酵素など、光合成装置の構成要素に似ていました。中でも目立ったのは、RuBisCO小サブユニット様タンパク質(RvRbcS-like)とRuBisCOアクチベーター様タンパク質(RvRca-like)でした。どちらも二酸化炭素を取り込む中心酵素RuBisCOの重要な補助因子に関連しています。これらの宿主遺伝子は餌を摂取した直後、クレプトプラストが再形成され長期利用の準備をされる時期に強く発現しました。
宿主タンパク質がクレプトプラスト内に入ることの証明
示唆に富む遺伝子配列を見つけるだけでは不十分で、タンパク質が実際にクレプトプラストの内部に到達する必要があります。カスタム抗体と蛍光顕微鏡を用いて、研究チームはRvRbcS-likeとRvRca-likeがどこに蓄積するかを追跡しました。RvRbcS-likeに小さな検出タグを付けたRapaza株を作製すると、そのシグナルはクレプトプラストやRuBisCO自身と重なっていることが示されました。RvRca-likeに対する抗体による同様のイメージングは、このタンパク質もクレプトプラスト内に濃縮していることを明らかにしました。タンパク質ゲル上では、両タンパク質とも短くなっているように見え、膜を越える際に特別な前端セグメントが切除されることと一致しました――これは通常の葉緑体へタンパク質を導く標的シグナルと同様です。
これらの借用された補助因子が重要な理由
これらの宿主タンパク質が本当に重要かをテストするため、著者らはCRISPRを用いたゲノム編集で各遺伝子をノックアウトしました。RvRbcS-likeを欠く細胞は成長不良になり、光合成能力を失い、エネルギーを蓄える粒子の生成が大幅に減り、通常よりはるかに早く死にました。藻類由来のRuBisCO大サブユニットの量も低下し、宿主による小サブユニットの代替がなければ酵素複合体が崩壊することを示唆します。RvRca-likeの除去はより穏やかな影響でしたが、それでも明瞭でした:初期の成長はほぼ正常でも、後期には光合成と炭素貯蔵が低下し、効率が徐々に失われることを示しました。これらの結果は、Rapazaが単に盗んだ葉緑体を放置しているのではなく、自らのタンパク質ツールキットでそれらを能動的に維持・再構築していることを示しています。
専用の配達タグと再構築された内部構造
Rapazaのクレプトプラスト標的と考えられるタンパク質の多くは、前端に長く低構造の「ヘッド」を共有し、しばしば膜貫通領域が予測されます。これらのヘッドの一つをルシフェラーゼ報告タンパク質に融合させることで、このセグメントだけで荷物タンパク質をクレプトプラストへ運べることが示されました。詳細な配列解析はこれらの標的領域のいくつかの類型を明らかにし、これらは関連藻類群が三重膜を持つ恒久的葉緑体へタンパク質を輸送する際に使うものと密接に似ています。特に注目すべき宿主タンパク質RvRbcS-likeは、RuBisCOに関連するドメインを四つ持ち、繰り返しモチーフに富む柔らかい尾部を備えています。この尾部はタンパク質の相分離を促すと考えられており、著者らはこの尾がクレプトプラスト内部の致密な炭素固定ハブであるピレノイドを複数のドロップレットに再編成し、それらが娘細胞に受け継がれるのを助けると提案しています。
複雑な細胞の物語にとっての意味
非専門家にとっての主要なメッセージは、Rapaza viridisが古代の細胞が自由生活する細菌を葉緑体のような恒久的部品に変えたときに起こったかもしれないことの、生きた可逆的なバージョンを示しているという点です。ここで宿主は急速に一時的な分子パートナーシップを構築します:宿主自身の遺伝子が重要な代替部品を供給し、特殊な配達タグに導かれて盗まれた葉緑体の内部構造まで再形成します。この「一時的分子キメラリズム」は、種間の短期的な統合であっても高度で精緻に調整され得ることを示しています。この系を研究することで、科学者は複雑な細胞が外来のエネルギー工場をどう制御・供給・安全に利用することを学んだか、そして新たな細胞小器官が進化の過程で今なおどのように出現し得るかを解き明かす強力なモデルを得られます。
引用: Kashiyama, Y., Maruyama, M., Nakazawa, M. et al. Transient molecular chimerism for exploiting xenogeneic organelles. Nat Commun 17, 2371 (2026). https://doi.org/10.1038/s41467-026-70516-x
キーワード: クレプトプラスティ, 葉緑体の進化, Rapaza viridis, 内共生, 細胞小器官の形成