Clear Sky Science · ja
DNAメチル化を欠く線虫Caenorhabditis elegansにおけるNuRD依存・非依存のMBD-2の二重ゲノム局在と遺伝子制御機能
小さな線虫が遺伝子制御のルールを書き換えるしくみ
私たちの細胞は、どの遺伝子をオンにするかオフにするかを記憶するために、DNAやタンパク質に化学的なタグを付けます。最も知られたタグの一つであるDNAメチル化は、一部の動物では失われていますが、それでも正常に成長し繁殖します。本論文は小さな線虫Caenorhabditis elegansがDNAメチル化なしでどのように遺伝子制御を行っているかを探り、予想外に柔軟な代替システムを明らかにしており、エピジェネティクスの考え方を再構築する可能性を示します。
失われたDNA標識が突きつける大きな疑問
哺乳類では5-メチルシトシン(5mC)と呼ばれる化学的標識が、発生過程やX染色体不活化、トランスポゾンの抑制などで遺伝子をサイレンシングするのに役立ちます。MBD2やMBD3と呼ばれるタンパク質はこれらの標識を読み取り、大型の蛋白質機械であるNuRD複合体を呼び寄せ、クロマチンの構造を再配列して遺伝子を抑制します。ところが興味深いことに、多くの無脊椎動物、例えばC. elegansやショウジョウバエは5mCとそれを付加する酵素を失っていますが、MBD2/3様タンパク質は保持しています。これは「DNAメチル化リーダー」を、もはやDNAメチル化を用いないゲノムでなぜ保持しているのか、そして代わりに何をしているのかという疑問を投げかけます。
線虫が失えない足場タンパク質
著者らはこのタンパク質の線虫版であるMBD-2に注目しました。哺乳類の同族と異なり、線虫MBD-2は5mCを認識する古典的なメチル結合ドメインを失っていますが、他のタンパク質と結合できる柔軟な領域やコイルドコイル領域を保持しています。MBD-2に蛍光タグを付けると、線虫のほぼすべての細胞の核に存在することが示され、広範な遺伝子制御の役割と整合しました。遺伝子工学を用いて、MBD-2を完全に欠失させた線虫や、NuRDとの接触を媒介するコイルドコイル領域だけを欠いた線虫を作成すると、いずれの変異でも深刻な異常が生じました:個体は小型で運動性が低下し、生殖器が奇形になり、ほとんど不妊になりました。これはDNAメチル化が存在しないにもかかわらず、MBD-2が正常な発生と生殖能力に不可欠であることを示しています。
メチル化のない世界でNuRD機構を再構築する
MBD-2が今でもNuRDと協働しているかを調べるため、著者らはタグ付きMBD-2を線虫抽出物から引き出し、質量分析で相互作用パートナーを同定しました。既知のNuRD構成要素の多くが存在し、MBD-2がこの複合体のコア足場として働いていることが確認され、哺乳類のMBD2/3と類似していることが示されました。コイルドコイル領域を削除すると、特にヌクレオソームを再構築するタンパク質との相互作用が多く失われました。同時に大規模なRNAシーケンス解析により、MBD-2変異体で全遺伝子の4分の1以上の発現が変化し、特に多くの遺伝子がオンになる方向に変化していることが明らかになりました。このパターンは、MBD-2がしばしばNuRDとともに不適切な遺伝子の発現を抑えるリプレッサーとして働く一方で、高発現遺伝子の一部の適正な活動を維持するのにも関与していることを示しています。
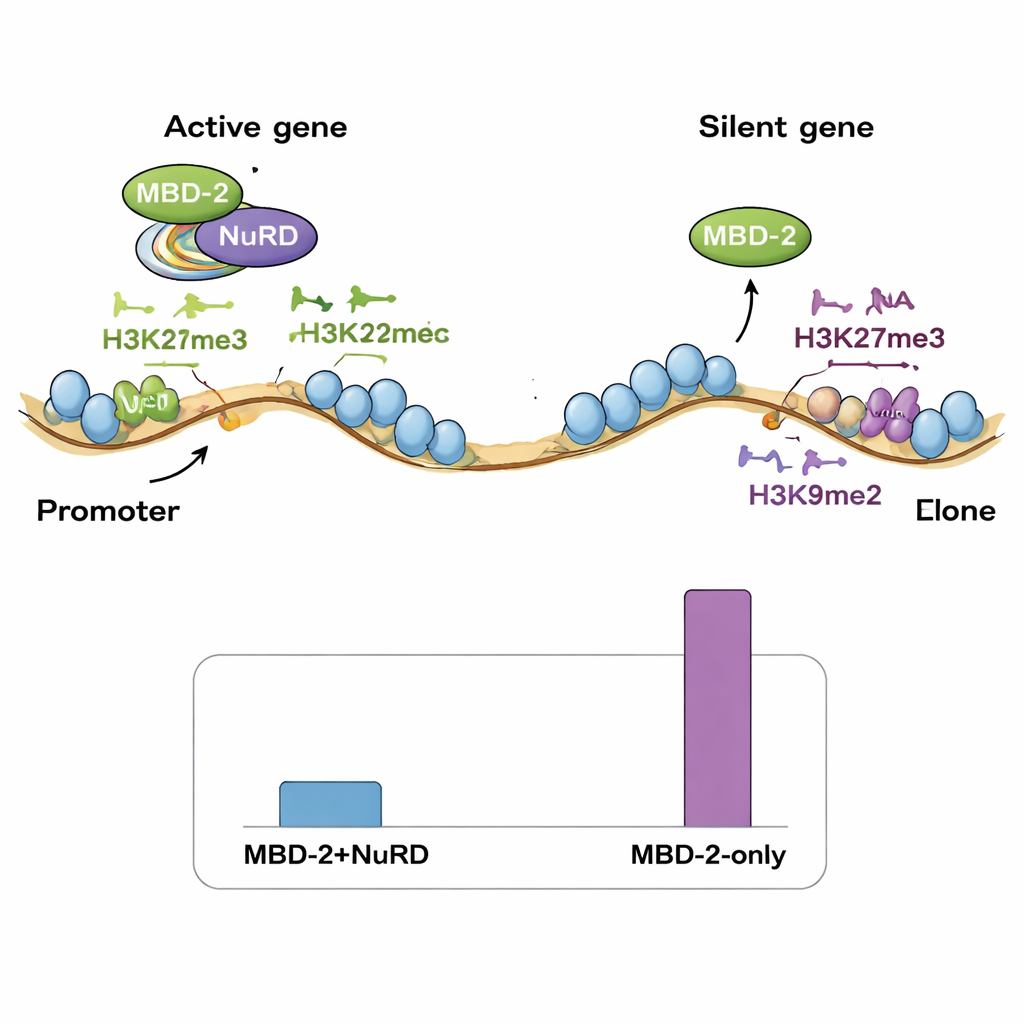
ゲノム上での二つの結合モード
次にチームはChIP-seqでMBD-2がゲノム上のどこに位置するかをマッピングし、既存のNuRD成分やさまざまなヒストン修飾の地図と比較しました。驚いたことに、MBD-2のサイトのごく一部だけがNuRDタンパク質や開いた活性クロマチンのマークと強く重なっていました。大部分のMBD-2結合部位は別の場所、すなわちH3K27me3やH3K9me2/3のような抑制的なヒストン標識が豊富な染色体の“腕”領域に位置していました。言い換えれば、MBD-2は二重の振る舞いを示します:少数のサイトではNuRDとともに移動して活性遺伝子を微調整しますが、多数のサイトでは既に沈黙している領域に独立して結合します。また、遺伝子内での結合位置も重要です—MBD-2が遺伝子の開始付近に集まると、そのタンパク質を失うとこれらの遺伝子が活性化する傾向があり、遺伝子本体の深部に結合する場合はMBD-2除去で発現が低下することが多いのです。
失われたDNAメチル化のための進化的バックアップ
著者らは、DNAメチル化を失った種では、特にH3K27me3といった抑制的なヒストン修飾が似た規制的ニッチを埋めるように拡大したのではないかと提案します。線虫のMBD-2は再利用され、DNA上の5mCを読み取る代わりに、特定のヒストン修飾で特徴付けられたクロマチン領域に結合するようになり、保存されたタンパク質相互作用ドメインを通じて依然としてNuRD複合体を固定しています。本研究は、古くからある遺伝子サイレンシングの道具立てが進化の中で書き換えられ得ることを示しています:DNA上の化学的標識が消えても、関連するタンパク質機構は代替のシグナルに切り替えて生き残ることができるのです。非専門家に向けた主要なメッセージは、遺伝子制御は驚くほど適応的であり、細胞は主要なエピジェネティック標識を失っても、他のクロマチンタッグやMBD-2のような多目的な足場タンパク質により数千の遺伝子の複雑な制御を維持できる、という点です。
引用: Tsui, H.N., Wong, C.Y.Y., Zheng, C. et al. Dual genomic localizations and gene regulatory functions of MBD-2 with and without NuRD in Caenorhabditis elegans which lacks DNA methylation. Nat Commun 17, 1875 (2026). https://doi.org/10.1038/s41467-026-68592-0
キーワード: エピジェネティクス, クロマチン, 遺伝子制御, C. elegans, ヒストン修飾