Clear Sky Science · ja
統合型マルチオミクスアトラスが明らかにするマウス胚原腸胚形成の時空間的制御ネットワークの階層
胚が体の設計図を組み立てる仕組み
ヒトを含むすべての哺乳類は、最初は小さな細胞の塊として始まり、短期間のうちに頭部、尾部、背側、腹側、内臓を備えた複雑な体へと急速に組織化されなければなりません。この劇的な形態変化は原腸形成と呼ばれる短い時期に起こります。ここで要約する論文は、マウス胚におけるこの過程を詳細かつ多層的にマップ化し、遺伝子、DNAのスイッチ(調節領域)、および化学的シグナルが時空間的に協調して細胞を将来の運命へ導く様子を示しています。
何千もの細胞が選択を下す様子を観察する
原腸形成の進行を追うために、研究者たちは早期から後期にわたる5つの近接した発生段階のマウス胚から35,000超の個々の細胞を解析しました。各細胞について、どの遺伝子が発現しているかだけでなく、どのDNA領域が開いてアクセス可能になっているかも測定しました。DNAが開いていることはその領域が調節スイッチとして機能し得ることを示します。これらのデータから、31の異なる細胞型を同定し、初期の可塑的な細胞がどのようにして外胚葉・中胚葉・内胚葉という3つの主要な胚葉へと段階的に分岐していくかをたどりました。また、新しい計算手法BioCREを開発し、遺伝子とそれを制御するDNA要素をより正確に結びつけることで、多くの重要な調節要素が制御する遺伝子から遠く離れた位置に存在することを明らかにしました。
実際の胚空間における3次元分子アトラスの構築
ほとんどの単一細胞解析法では各細胞の元の胚内位置情報が失われますが、位置はパターニングを理解するうえで重要です。研究チームは既存のマウス胚における遺伝子活動の三次元マップに自らの単一細胞データを整列させることでこの問題を克服しました。その結果得られたST-MAGICは、“デジタル胚”であり、各微小領域に存在するであろう細胞型、発現している遺伝子、および周辺DNAのアクセス可能性が注釈されています。このアトラスは、例えば将来心臓や筋肉、支持組織になるさまざまな中胚葉サブタイプが異なる領域で出現する様子や、Otx2のように広く発現する遺伝子が胚の外側(胚盤葉上層)と内側(内臓内胚葉)で異なる制御要素を使うことを示しています。
左右の非対称性が生まれるとき
体の設計図の特筆すべき特徴の一つは左右が同一ではないことです—心臓がわずかに左に寄っていることを思い浮かべてください。著者らは空間アトラスを用いて左右差が最初に現れる側胚葉に焦点を当てました。そこで、各側でどの細胞型が偏在するか、どのDNA領域がより開いているかに関して微妙だが一貫した非対称性を見出しました。右側ではBMPと呼ばれる成長シグナル経路に関連する領域がよりアクセス可能であり、左側では節形成や心構造形成に必要な遺伝子に関連する領域が優勢でした。Lefty2を制御する新たに発見された要素を含め、これらのDNA要素のいくつかは遺伝子発現の目に見える差異よりも前に開き始め、クロマチンの早期“プライミング”が胚の各側がシグナルを異なって解釈する準備をすることを示唆しています。
体軸中央線を導く因子のリレー
研究は次に軸性中内胚葉(axial mesendoderm)に注目します。これは脊索を形成する細胞群で、脊索は体の正中線に沿う棒状構造として脊柱や神経系の組織化を助けます。ST-MAGICと拡張版のST-MAGIC (+)を用いて、著者らはこれらの細胞が前原始条(anterior primitive streak)からどのように生じ、ノード細胞(左右感知に重要な繊毛構造を作る)と前軸性中内胚葉(正中線組織に寄与する)という二つの枝に分かれるかを追跡しました。彼らは転写因子の階層的なリレーを明らかにしました。EOMESのような初期因子やFOXA2、LHX1といった中間段階の因子がまず重要なDNA領域を開き、WNTやNODALなど主要なシグナルへの応答性を確立します。後期の“終末”因子、例えばNOTO、SOX9、そして今回新たに関与が示唆されたPOU6F1は、繊毛や細胞外マトリックスに必要な専門的な遺伝子プログラムをオンにします。
シグナル、クロマチン、そして後期に働く専門因子
WNTおよびNODALのシグナル伝達因子がDNAへ結合する位置に関する公開データを統合することで、著者らは胚のシグナルに対する応答性がシグナル源自体が移動するより前に空間的に変化することを示しました。軸性中内胚葉へとプライムされる領域では、NODALおよびWNTに応答するDNA部位が早期に開き、多くはFOXA2、Zfp281、その他の制御因子のモチーフを含んでおり、協調的な制御を示唆します。マウスにおける後期因子NOTOおよびPOU6F1の実験的欠失は、ノード特異的および繊毛関連遺伝子の発現を乱し、ノード繊毛を短縮させましたが、基盤となる開放されたDNAのランドスケープは大部分が維持されていました。これは、初期の因子がエピジェネティックな基盤を築き、後期の因子は主にクロマチンを再編成することなく遺伝子発現を精緻化する役割を果たすことを示しています。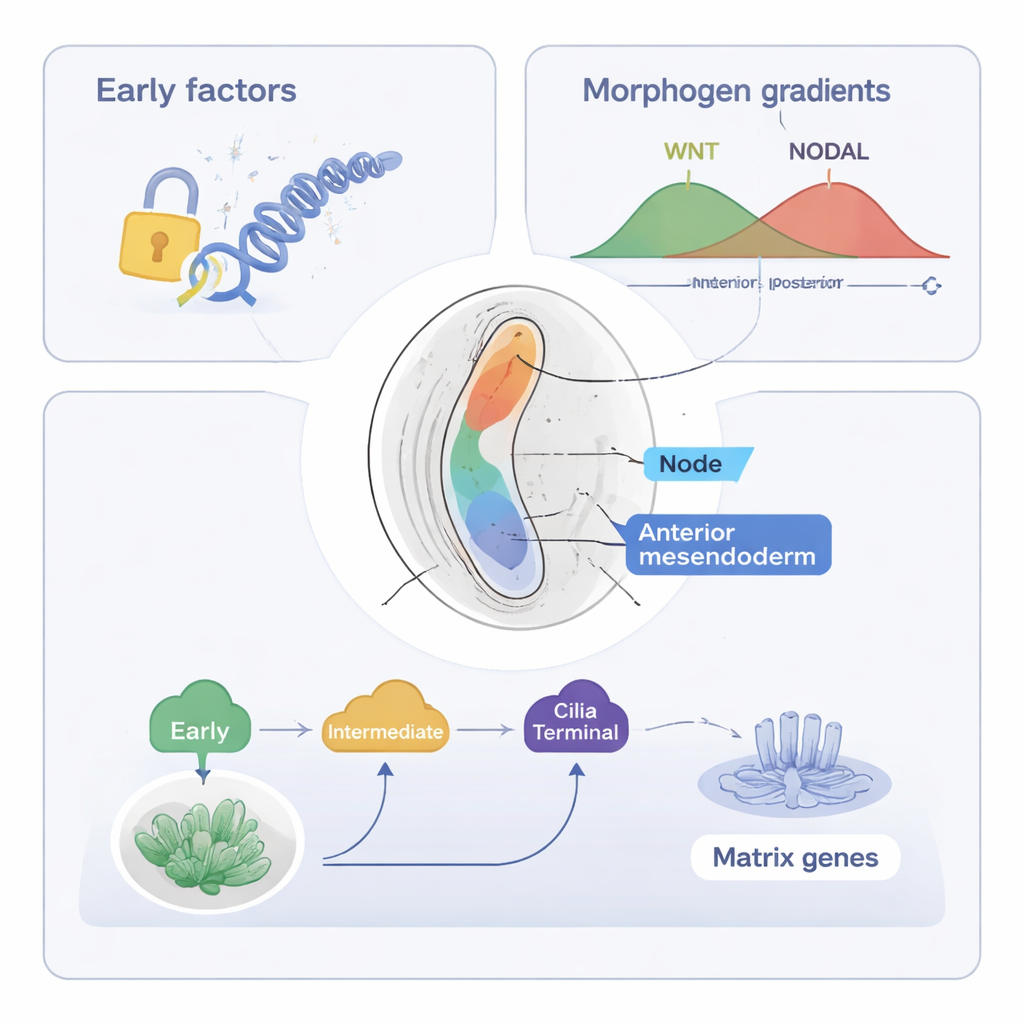
なぜ発生理解に重要なのか
専門家でない読者にとって、この研究は胚が体の設計図をどのように敷設するかの高解像度な“配線図”を構築したものとして見ることができます。著者らは、原腸形成期の細胞運命決定は存在するシグナルだけで決まるのではなく、いつどこでDNAスイッチが開かれるか、そしてどの転写因子が順序を追って作用するかによっても支配されることを示しました。彼らのST-MAGICおよびST-MAGIC (+)アトラスは、時空間にわたるこれらの関係を探るためのリソースを提供し、先天性欠損の研究、幹細胞ベースの胚モデル、および最終的にはヒト発生の側面に関する研究に貢献し得る枠組みを提示します。
引用: Yang, X., Xie, B., Shen, P. et al. Integrated multi-omic atlas reveals the hierarchy of spatiotemporal regulatory networks of mouse gastrulation. Nat Commun 17, 1572 (2026). https://doi.org/10.1038/s41467-026-68291-w
キーワード: 原腸形成, 遺伝子制御ネットワーク, 単一細胞マルチオミクス, 胚のパターニング, マウス発生