Clear Sky Science · it
Ruoli distinti dei sottotipi della corteccia strato 5 nell’apprendimento associativo
Come il cervello impara che un tocco predice una ricompensa
Immagina di imparare che un leggero tocco sul polso significa che arriverà il dessert. Il cervello deve collegare un semplice tocco a una ricompensa futura. Questo studio esplora il cervello del topo per vedere come due diversi tipi di cellule nervose lavorino insieme per formare questi collegamenti, rivelando come l’apprendimento quotidiano sia diviso tra “cosa è successo” e “cosa significa per me”. 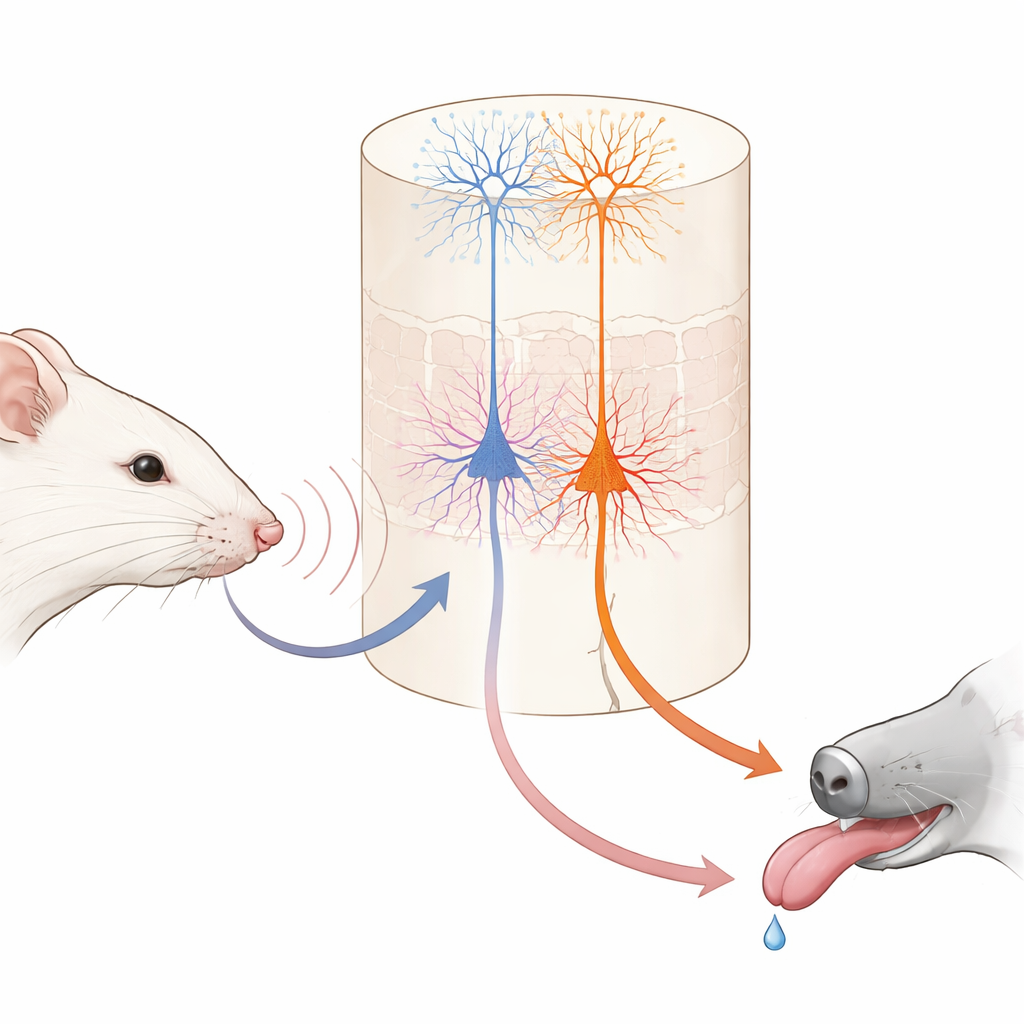
Un semplice gioco dei baffi per studiare l’apprendimento
I ricercatori hanno addestrato topi con la testa fissata a un compito semplice usando i loro baffi facciali. In ogni prova, un singolo baffo veniva vibrato a una di due velocità. Una vibrazione era seguita, dopo un breve intervallo, da una minutissima goccia d’acqua; l’altra non veniva mai ricompensata. All’inizio i topi leccavano in anticipazione durante entrambi i tipi di prova, non sapendo ancora quale sensazione sul baffo segnalasse l’acqua. Nel corso di alcuni giorni hanno gradualmente imparato a leccare presto solo quando si verificava la vibrazione “buona” e a trattenersi quando appariva quella non ricompensata. Quando gli scienziati hanno temporaneamente spento l’area primaria del tatto del cervello durante l’allenamento, questo apprendimento è in gran parte scomparso, dimostrando che questa regione sensoriale è importante per costruire l’associazione, anche se i topi esperti poi potevano eseguire il compito senza di essa.
Due tipi di cellule di output con compiti molto diversi
All’interno di quest’area tattilе, uno strato profondo noto come strato 5 contiene due principali tipi di cellule nervose di output. Un gruppo, chiamato qui cellule IT, invia segnali ad altre regioni corticali di entrambi gli emisferi cerebrali. L’altro gruppo, cellule ET, invia segnali prevalentemente verso strutture sottocorticali coinvolte nell’azione e nella ricompensa. Utilizzando topi geneticamente modificati e imaging a due fotoni ad alta risoluzione, gli autori hanno potuto monitorare selettivamente l’attività di ciascun tipo cellulare attraverso le punte dei loro rami lunghi e simili ad alberi. Prima dell’apprendimento, le cellule IT rispondevano già in modo forte e affidabile alle vibrazioni dei baffi, e la loro attività combinata poteva distinguere con precisione le due velocità di vibrazione. Le cellule ET, al contrario, rispondevano in modo più debole e meno coerente agli stimoli, offrendo solo una lettura sfocata di quale vibrazione fosse avvenuta.
Sensazioni stabili contro aspettative in crescita
Man mano che i topi imparavano, le cellule IT si comportavano come affidabili cronisti. Le loro risposte restavano strettamente legate al momento del movimento del baffo e cambiavano poco da un giorno all’altro. Continuavano a codificare quale vibrazione fosse avvenuta, indipendentemente dal fatto che predicesse o meno una ricompensa. Le cellule ET, invece, trasformavano il loro comportamento. Invece di limitarsi a scattare all’insorgere dello stimolo, la loro attività aumentava gradualmente durante la vibrazione e il breve intervallo, raggiungendo un picco intorno al tempo previsto per la somministrazione dell’acqua. Questo aumento cresceva in parallelo con le leccate anticipatorie degli animali e diventava più abile nel predire se una prova sarebbe terminata con una leccata piuttosto che nel riportare esattamente lo stimolo. Singole cellule ET entravano e uscivano dal gruppo attivo nel corso dei giorni, ma a livello di popolazione il modello diventava più coerente, suggerendo un codice flessibile ma convergente per l’aspettativa di ricompensa. 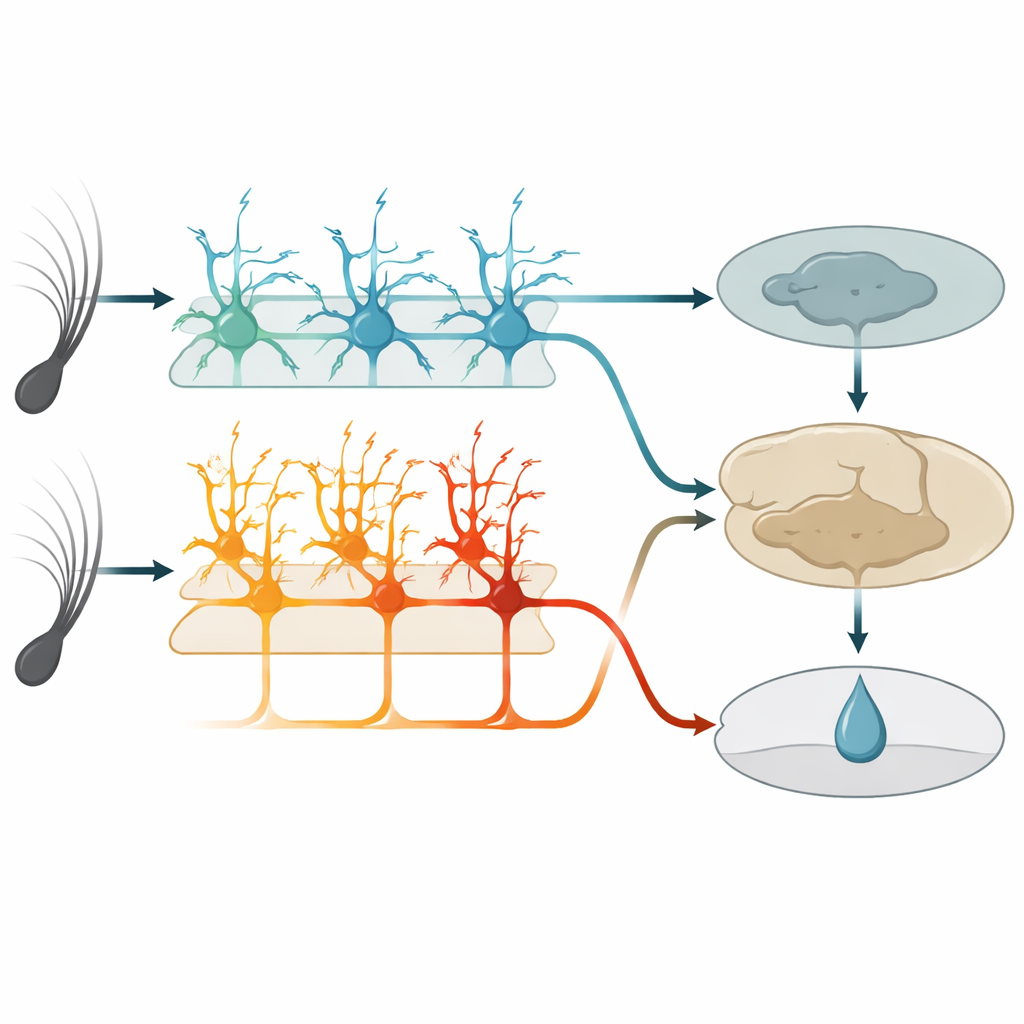
Spegnere ciascun tipo cellulare rivela una divisione del lavoro
Per testare la funzione, il team ha usato strumenti chemogenetici per smorzare selettivamente le cellule IT o ET durante l’addestramento. Quando le cellule IT venivano silenziate, i topi mostravano in generale meno leccate anticipatorie e non riuscivano a costruire una chiara differenza tra vibrazioni ricompensate e non ricompensate. Quando le cellule ET venivano silenziate, i topi facevano l’opposto: leccavano troppo in entrambe le vibrazioni, soprattutto quella non ricompensata, e faticavano a affinare il loro comportamento nonostante continuassero a leccare vigorosamente. Spegnere uno dei due gruppi dopo che il compito era stato padroneggiato non danneggiava più la prestazione, implicando che una volta che altre regioni cerebrali hanno immagazzinato l’associazione, quest’area sensoriale e i suoi output dello strato 5 diventano meno critici per eseguire la risposta appresa.
Un modello di apprendimento che rispecchia il comportamento cerebrale
Gli autori hanno costruito un modello computazionale in stile apprendimento per rinforzo per interpretare questi risultati. Nel modello, una rete simile alle IT fornisce rappresentazioni sensoriali stabili che aiutano a stimare il “valore” di ciascuno stimolo—quanto è probabile che sia seguito da una ricompensa. Un percorso simile alle ET trasmette questo valore previsto a un circuito downstream che lo confronta con la ricompensa effettiva, generando un errore di predizione che aggiusta le stime di valore future. Bloccare i percorsi IT o ET nel modello ha riprodotto i distinti fallimenti dell’apprendimento osservati nei topi: senza input simile alle IT, l’apprendimento era lento e debole per entrambi gli stimoli; senza output simile alle ET, l’apprendimento iniziale avveniva ma il sistema non riusciva a ridurre adeguatamente le risposte al segnale non ricompensato. Il modello ha inoltre catturato come, col tempo, regioni non sensoriali potessero subentrare nella performance, coerentemente con gli esperimenti.
Cosa significa per l’apprendimento quotidiano
In termini semplici, questo studio suggerisce che quando impariamo che un particolare aspetto visivo, sonoro o tattile predice qualcosa di buono o cattivo, diversi gruppi di neuroni corticali profondi condividono il lavoro. Un gruppo conserva un resoconto fedele di “cosa è successo” nel mondo, mentre un altro gradualmente comincia a segnalare “cosa mi aspetto che accada dopo” e contribuisce a perfezionare il comportamento confrontando quelle aspettative con la realtà. Insieme formano un ponte tra la sensazione grezza e azioni flessibili basate sull’esperienza, offrendo un quadro più chiaro di come il cervello supporta le abitudini, l’apprendimento da ricompense e, possibilmente, come questi processi possono andare storti nelle malattie.
Citazione: Moberg, S., Garibbo, M., Mazo, C. et al. Distinct roles of cortical layer 5 subtypes in associative learning. Nat Commun 17, 2648 (2026). https://doi.org/10.1038/s41467-026-68307-5
Parole chiave: apprendimento associativo, corteccia somatosensoriale, neuroni dello strato 5, aspettativa di ricompensa, apprendimento per rinforzo