Clear Sky Science · he
מבנה ותפקוד של הסופרקומפלקס החושי לחומצות אמינו SEAC–EGOC בשמרים
איך תאים מחליטים מתי לגדול
כל תא צריך כל הזמן להכריע האם הוא יכול להרשות לעצמו לגדול או שמוטב לשמר משאבים. ההחלטה הזאת תלויה במידה רבה בחומצות אמינו, אבני הבניין של חלבונים ומקור מרכזי לחנקן. המאמר חושף, בפירוט מבני מרשים, כיצד תאי שמרים חשים זמינות חומצות אמינו דרך מחסום מולקולרי ענק ומשתמשים במידע הזה כדי להפעיל או לכבות בורר גדילה מרכזי בשם TORC1. מאחר שמעגלים מקבילים שמורים גם בתאים אנושיים והם מקושרים לסרטן ולמטבוליזם, התובנות משמרים מספקות חלון להבנת הדרך שבה תאי גוףנו שוקלים האם התנאים מתאימים לצמיחה.
צומת מתוחכם במרכז המיחזור של התא
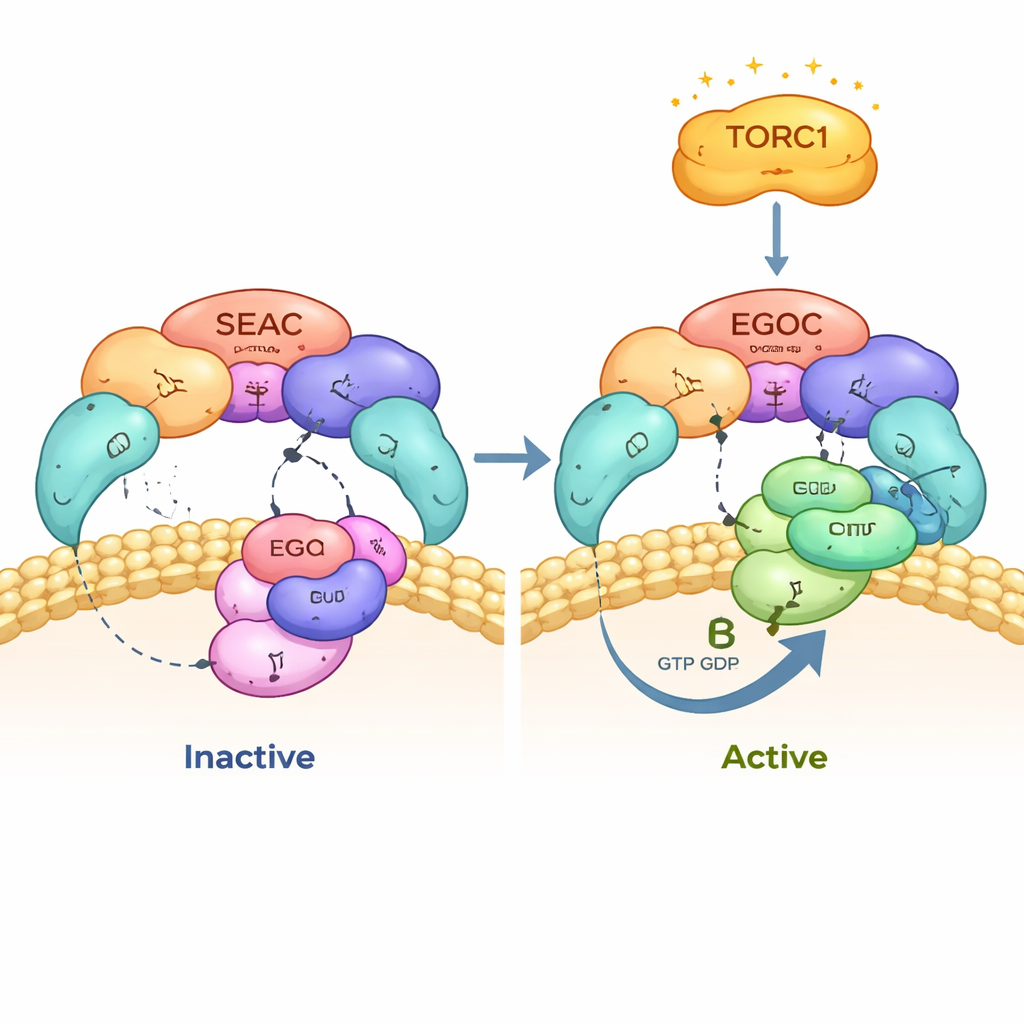
בשמרים, קומפלקס מרכזי לקבלת החלטות הנקרא SEAC יושב על ממברנת הואקואול, האברון שממלא תפקיד של מרכז מיחזור בתא. שם הוא פוגש קומפלקס נוסף, EGOC, שמעביר מידע על רמות חומצות האמינו. יחד הם מזינים אותות אל TORC1, מתג הגדילה הראשי. ל-SEAC יש שתי חצאים תפקודיות: SEACIT, שיכולה לכבות את TORC1, ו-SEACAT, שאמורה מאז ומתמיד להחזיק את SEACIT במעקב. החוקרים השתמשו במיקרוסקופיה קריואלקטרונית ברזולוציה גבוהה כדי לראות את ה-SEAC במלואו קשור ל-EGOC, ולתעד תמונת רגע של הצומת המולקולרי בדיוק במקום שבו מתבצע ההשידור, על פני השטח של הוואקואול. 
מבט מקרוב על האדריכלות המולקולרית
המבנה חושף את ה-SEAC כגרעין גדול העשוי תת-יחידות SEACAT, שממנו מתארכות שתי "כנפיים" גמישות של SEACIT. כל כנף יכולה לקשור קומפלקס EGOC אחד, כלומר SEAC יחיד מסוגל לקלוט שתי כניסות איתות בו־זמנית. באופן בולט, EGOC מתחבר רק לכנפיים ולא לגרעין, וצורת ה-SEAC כמעט ואינה משתנה בנוכחות EGOC. זה מצביע על כך ש-SEACAT אינו חוסם פיזית את האתר הפעיל של SEACIT ואינו מונע מ-EGOC להיקשר. האוריינטציה של הקומפלקס על ממברנת הוואקואול גם מציבה את גרעין ה-SEAC הפונה לציטופלזמה, שם הוא יכול לתקשר עם רגולטורים נוספים, בעוד שהכנפיים שוכנות קרוב לממברנה, במיקום אידיאלי ליצירת מגע עם EGOC ושאר שותפים מעוגני ממברנה.
כיצד SEAC בוחר את האות הנכון
המחקר זומזם על המגעים שבין SEAC ל-EGOC ומצא ש-SEAC מזהה רק את הגרסה "הפעילה" של EGOC, שבה אחת מחלבוני ה-GTPase הקטנים שלו, Gtr1, נושאת נוקלאוטיד בדומה ל-GTP, בעוד ששותפו Gtr2 מחזיק GDP. SEACIT מספקת "אצבע" קטליטית שנכנסת לכיס הנוקלאוטיד של Gtr1 ומזרזת את פירוק ה-GTP, והופכת את EGOC ממצב פעיל לבלתי פעיל. מפות קריו-EM מפורטות וניסויי מוטציות מראים שאצבע קטליטית זו ושאר חסיד עזר סמוך חיוניים לאינטראקציה ולקריאת מצב הנוקלאוטיד כראוי. כאשר החוקרים שינו חומצות מפתח אלו, ה-SEAC חדל ממיקומו התקין על הוואקואול, ו-EGOC לא יכל להיקשר בצורה פרודוקטיבית.
בקרה מהירה לעומת איטית על מתג הגדילה
כדי להבין מה משמעות הממצאים בתא החי, החוקרים עקבו אחרי פעילות TORC1 לאורך זמן כאשר חומצות האמינו הוסרו ואחר כך הוחזרו. בתאים תקינים, TORC1 נכבה במהירות בתוך מספר דקות של חשבון חנקן ואז הופך לחזור ולהידלק במהירות כשחומצות האמינו חוזרות. כאשר האצבע הקטליטית ב-SEACIT נעצבה, או שתתי-היחידות של SEACIT הוסרו, TORC1 כמעט ולא הגיב בחלון הזמן המוקדם הזה. כך היה גם כאשר הוסרו גם Gtr1 וגם Gtr2, מה שמראה שצמד SEAC–EGOC הוא המסלול הדומיננטי והמהיר בשליטת חומצות האמינו על TORC1. עם זאת, בזמנים מאוחרים יותר—בערך חצי שעה—פעילות TORC1 הותאמה באיטיות גם במוטנטים אלה, דבר המעיד שקיים מסלול נפרד ואיטי יותר אל TORC1, עצמאי ממערכת ה-GTPase הזו, שפועל גם הוא בתאים.
תפקיד מפתיע של תת-יחידה מרוחקת
ממצא מעניין במיוחד נוגע ל-Sea2, מרכיב של SEACAT עם תחום "מדחף" גמיש שנוטה החוצה מהגרעין. הסרה של Sea2, או רק של המדחף שלה, נתנה ליקויים באיתות הדומים כמעט בדיוק לאלה שנצפו כאשר הפעילות הקטליטית של SEAC הושבתה: TORC1 הגיב באיטיות להחרמה ולהחזרת חומצות האמינו. עם זאת, Sea2 יושבת רחוק מאתר הקטליזה ואינה נוגעת ישירות בכנף SEACIT, מה שמקשה על הסבר המבוסס על חסימה פשוטה. ניסויים גנטיים שמשלבים חוסר Sea2 עם וריאנט SEAC חסר פעילות קטליטית הציעו שכאשר פעילות ה-GAP נעולה לגמרי במצב פעיל או בלתי פעיל, שניהם מופרעים בתגובה המהירה, בעוד שנחוצה ויסות ביניים. המחברים מציעים שמדחף Sea2 משמש כפלטפורמת עגינה לגורם לא ידוע שממתן את פעילות SEACIT, ומאפשר לתאים לכוונן את TORC1 במקום לכבות אותו באופן תמידי.
מה משמעות הדבר להחלטות גידול תאיות
בסך הכול, המחקר משכתב את ההבנה של SEAC — לא כשתי יחידות מנוגדות אלא כמכונה אחת משולבת שבה הכנף הקטליטית (SEACIT) והגרעין המבני (SEACAT) פועלות יחד כדי לחוש חומצות אמינו ולהתאים במהירות את מתג הגדילה TORC1. הפעולה האנזימטית של SEACIT על EGOC מספקת בקרה מהירה ומדויקת, בעוד שמדחף Sea2 כנראה מגייס רגולטורים שמודלים פעילות זו. במקביל, מסלולים איטיים יותר ומקבילים מבטיחים שפעילות TORC1 תסתנכרן בסופו של דבר עם מצב המזון גם כאשר החיישן הראשי מופרע. מאחר שקומפלקסים קרובים מבצעים תפקיד דומה בתאים אנושיים, התובנות משמרים מאירות כיצד תאי הגוף שלנו מאזנים בין גידול לזמינות מזון, עם השלכות להבנת מחלות שבהן איתות TORC1 משתבש.
ציטוט: Tafur, L., Bonadei, L., Zheng, Y. et al. Structure and function of the yeast amino acid-sensing SEAC–EGOC supercomplex. Nat Struct Mol Biol 33, 488–498 (2026). https://doi.org/10.1038/s41594-026-01746-2
מילות מפתח: חישה של חומצות אמינו, אותות TORC1, קומפלקס SEAC/GATOR, בקרת גידול תאים, מיקרוסקופיה קריואלקטרונית