Clear Sky Science · he
פוקוזילטרנספראז צמחי FUT11 מעוות את מקבל הסוכר כדי לקטלז באמצעות מנגנון בינוני חמצוני אוקסוקארבוקסיל זמני
איך צמחים מחדדים את שיוכי הסוכרים על החלבונים שלהם
חלבונים בתאינו, וכמו גם בצמחים, לעיתים קרובות מעוטרים בשרשראות סוכר מורכבות המשמשות כמעין ברקודים, המנחות כיצד החלבונים מתקפלים, כמה זמן הם מתמידים ובאילו שותפים יכולים הם להיקשר. המחקר הזה מתמקד באנזים צמחי בשם FUT11 שמוסיף סוכר קטן, פוקוזה, לשרשראות האלה. על-ידי גילוי מדויק של אופן פעולת FUT11 ברמת האטום, המחברים מגלים טריק מפתיע: האנזים לכאורה מעוקם באופן זמני את סוכר המקבל כדי לקדם שלב כימי קשה. הבנת התהליך חשובה לא רק לביולוגיה בסיסית, אלא גם לתכנון תרופות מיוצרות-צמח שמקטינות את הסיכון לעורר תגובות חיסוניות בלתי רצויות בבני אדם.
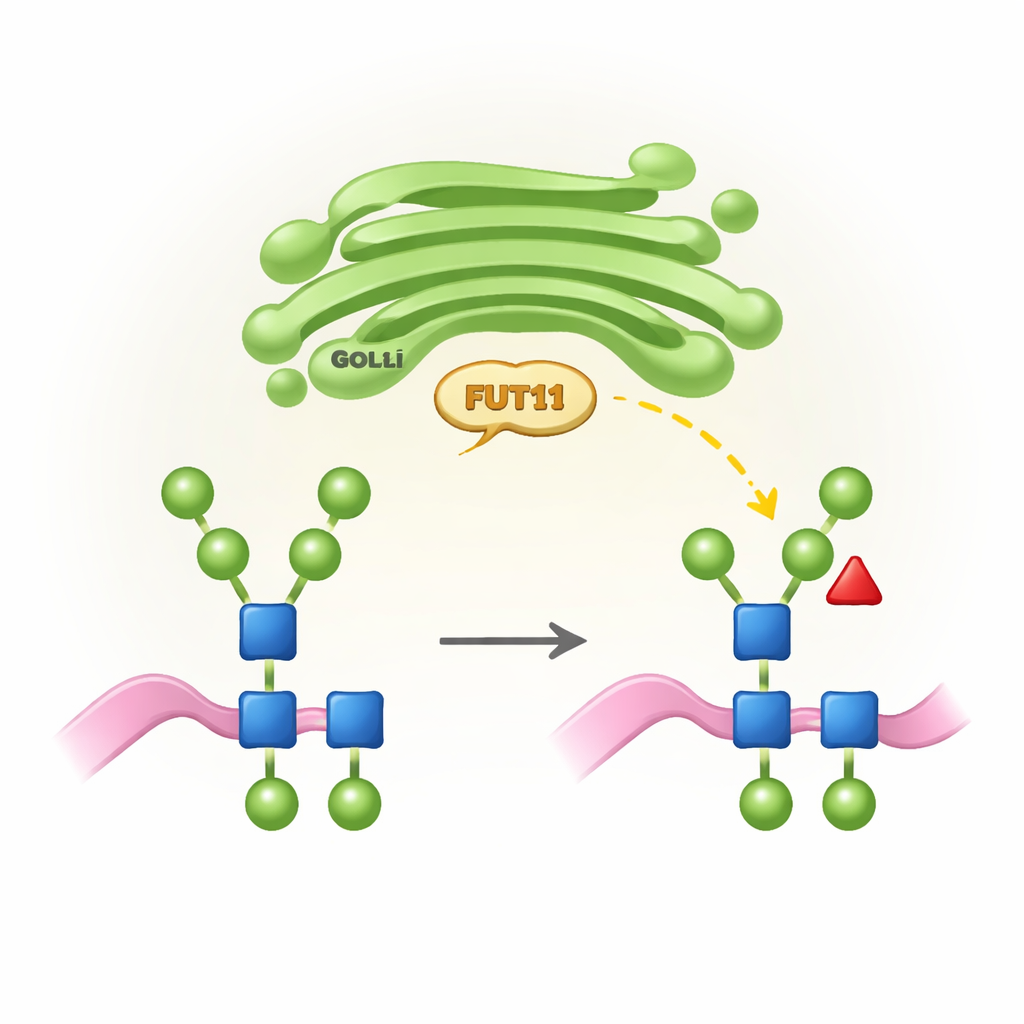
תגיות סוכר ספציפיות לצמחים ולמה הן חשובות
כל האורגניזמים המתקדמים משתמשים בתהליך שנקרא N-גליקוזילציה כדי לחבר שרשראות סוכר (N-גליקנים) לנקודות מסוימות על חלבונים. בבעלי חיים, שרשראות אלו מעובדות במערך גולג'י של התא למבנים מורכבים שלעיתים מסתיימים בחומצות סיאליות ונושאים פוקוזה ליבתית במיקום ספציפי אחד. צמחים, לעומת זאת, פועלים לפי "כלל עיצוב" שונה: ה-N-גליקנים שלהם בדרך-כלל חסרים חומצות סיאליות אך נושאים פוקוזה ליבתית במיקום אחר (הנקרא b1,3) וכן סוכר קסילוזה נוסף. התכונות הספציפיות האלה של הצמחים חיוניות לגידול ולפוריות תקינים, אבל הן יכולות להיתפס כזרות על-ידי המערכת החיסונית האנושית. FUT11 הוא אחד האנזימים המרכזיים בצמחים שמתקין את הפוקוזה הליבתית הזו, ופעילותו מעצבת הן את התפתחות הצמח והן את האופן שבו חלבונים תרופתיים מיוצרים-צמח ייתפסו בגופנו.
מיפוי מקומות שבהם FUT11 יכול לפעול על ענפים סוכריים מורכבים
כדי להבין מה FUT11 מזהה, החוקרים בחנו את האנזים על מיקרואריי של 144 מבני N-גליקן שונים. הם מצאו כי FUT11 סלקטיבי למדי לגבי אחד מהענפים של עץ הסוכר — זרוע הנקראת b1,3 — שצריכה לשאת יחידת בנייה מסוימת (N-אצטילגלוקוזאמין קיצוני, או GlcNAc) כדי שהתגובה תתקדם. במקביל, FUT11 סובלני לקישוטים אחרים: הוא עדיין פועל כאשר המנוזה המרכזית נושאת קסילוזה ייחודית לצמחים, ואף כאשר פוקוזה נוספת כבר הוכנסה במיקום היונקתי של בעלי חיים בליבה. הזרוע הנגדית (b1,6) חופשית ברובה לסביבה ועושה מגעים חלשים בלבד עם האנזים, מה שמסביר מדוע FUT11 יכול להתמודד עם מגוון רחב של שינויים שם. העדפות הקישור הללו מסייעות להסביר מדוע צמחים מייצרים סט אופייני של מבני N-גליקנים ומראות כיצד ניתן להשתמש ב-FUT11 או להימנע ממנו בהנדסה של תאי צמח לביוטכנולוגיה.
עיצוב מבני ייחודי לתפיסת וסידור הסוכרים
באמצעות גבישיות קרני רנטגן, הצוות פתר את המבנה התלת-ממדי של FUT11 קשור לתורם הסוכר שלו (GDP-פוקוזה) ולמקבל N-גליקן. האנזים מציג ארכיטקטורת "GT-B" בעלת שתי אונות: אונה אחת תומכת ב-GDP-פוקוזה, בעוד שאונה המקבל — מורכבת באופן בלתי רגיל וכוללת תת-דומיין N-טרמינלי ספציפי לצמחים — עוטפת את ה-N-גליקן. תת-הדומיין הנוסף הזה, המקושר לשאר האנזים באמצעות גשרים די-סולפידיים, מעגן את החלק המרכזי של הגליקן ועוזר להציג את יחידת ה-GlcNAc הריאקטיבית מול התורם. שינוי חומצות אמינו מפתח גיבש את תפקידיהן: החלפת גלוטמט אחד (Glu158) השביתה את הפעילות, בעוד ששינוי שתי שיירים קרובים החליש באופן ניכר את הפוקוזילציה בתאים אנושיים מהונדסים. התוצאות מקשרות בין תמונות מבניות לפונקציה תאית ממשית, ומראות בדיוק אילו חלקי החלבון חיוניים לקשירה ולקטליזה.
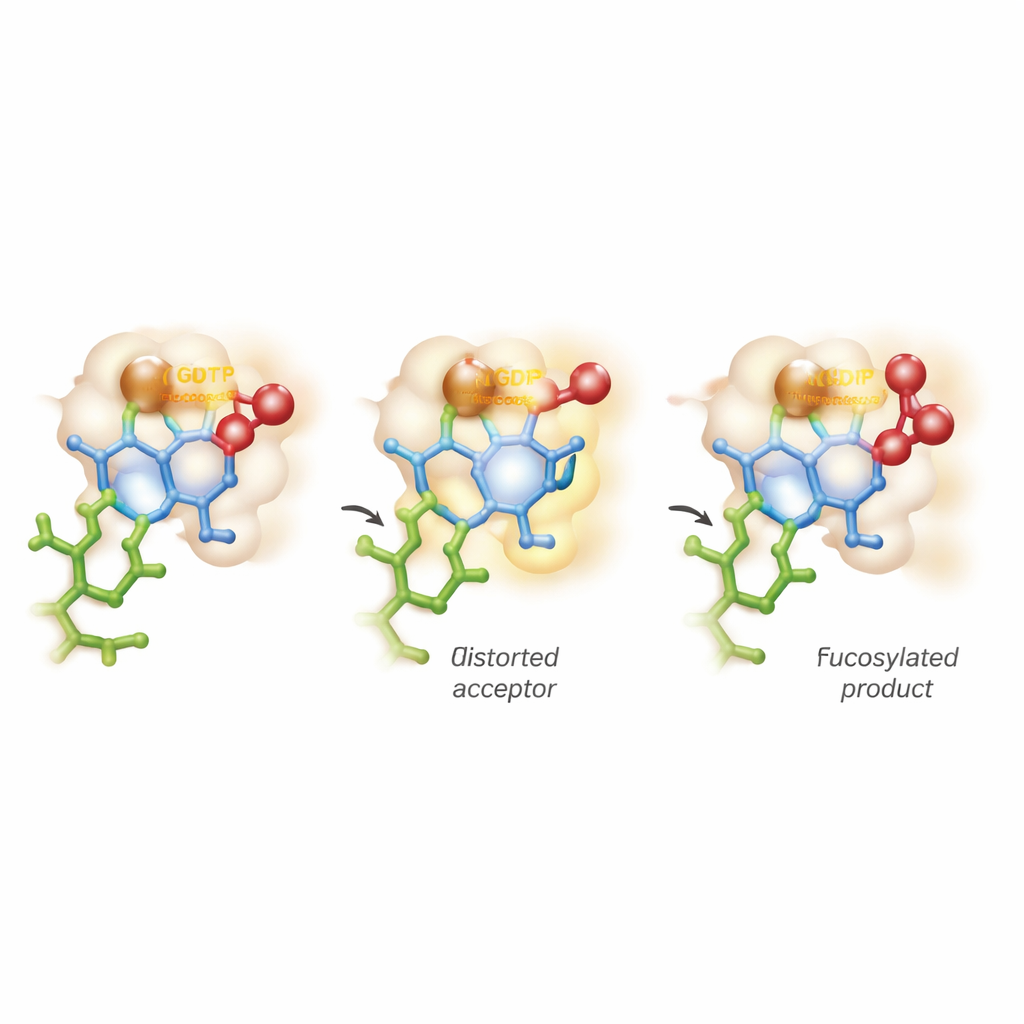
כיפוף הסוכר והימצאות בין שני מסלולי תגובה
התובנה הבולטת ביותר נובעת מדימות ממוחשבת מתקדם שמשלבת מכניקת קוונטים ודינמיקת מולקולות. התפיסות המסורתיות מניחות שמבנה הכיסא היציב של סוכר המקבל נשמר בזמן שהאנזים רק מפעיל את התורם. כאן, FUT11 פועל באגרסיביות רבה יותר: אינטראקציות המונעות על-ידי Glu158 מעוותות זמנית את טבעת ה-GlcNAc הפנימית לצורה פחות נוחה ו"מפוקפצת". בתנוחה המותחת הזו, קבוצת ההידרוקסיל הקריטית מיושרת בצורה מושלמת לתקוף את תורם הפוקוזה. החישובים מגלים שכאשר הקשר הכימי אל ה-GDP נשבר, מופיע מצב קצר-חיי ומטען חיובי הדומה לאוקסוקארבניום על הסוכר, לפני שהקשר החדש למקבל מתבצע במלואו. משמעות הדבר היא שהתגובה אינה מתיישבת באופן נקי בקטגוריות ה-SN1 או SN2 המוכרות, אלא מתקדמת על רצף, בצורה אסינכרונית ובעצם בצעדים כמעט נפרדים.
גמישות חבויה והדים אבולוציוניים
בהשוואה בין FUT11 לבין אנזים קרוב של בני אדם, FUT9, המחברים מצאו כי האנזים הצמחי יכול גם, אם כי בחולשה, לשנות סוג סוכר שונה (LacNAc) כדי ליצור מבנה הידוע כ-Lewis X. פעילות צד זו כנראה אינה חשובה ביולוגית בצמחים, שבהם Lewis X אינו נצפה באופן רגיל, אך היא מדגישה כיצד שלדות חלבוניות דומות יכולות לעבור מיחזור במהלך האבולוציה כדי לפעול בהקשרים סוכריים שונים. הממצאים מציעים כי ל-FUT11 ולקרוביו יש מסגרת מודולרית לזיהוי N-גליקנים, עם התאמות עדינות שמזיזות את הספציפיות בין ליבות צמחיות לבין אנטנות חיות.
למה הטריק של הכיפוף חשוב למדע ולרפואה
במבט כולל, העבודה מראה כי FUT11 עושה יותר מאשר לקרב בין שני שותפי סוכר: הוא מעקל באופן אקטיבי את סוכר המקבל לפוזה ריאקטיבית ומכוון את הכימיה דרך בינוני טעון רגעי. עבור הקורא הכללי, משמעות הדבר היא שהסוכר על החלבון אינו אתר עיגון קשיח אלא שותף גמיש שהאנזים מעצב לפי צרכיו. התפיסה החדשה הזו של "קטליזה קונפורמציונלית" מסבירה כיצד אנזימים משיגים גם מהירות וגם סלקטיביות ומספקת תבנית לעיצוב מסלולי גליקוזילציה. במונחים מעשיים, הידע המדויק כיצד FUT11 מזהה ומעבד N-גליקנים בצמחים יכול להנחות הנדסת צמחים ומערכות ייצור מבוססות צמחים כדי למזער דפוסי סוכר שמעוררים חיסון בבן אדם, או כדי ליצור גליקופroteינים מעוצבים עם תכונות ביולוגיות מותאמות.
ציטוט: Taleb, V., Sanz-Martínez, I., Serna, S. et al. Plant fucosyltransferase FUT11 distorts the sugar acceptor to catalyze via a transient oxocarbenium intermediate mechanism. Nat Commun 17, 1960 (2026). https://doi.org/10.1038/s41467-026-68786-6
מילות מפתח: גליקוזילציה בצמחים, פוקוזילטרנספראז FUT11, אנ-גליקנים, מנגנון אנזימטי, הנדסת גליקו