Clear Sky Science · fr
Prolifération transitoire par contrôle réversible de YAP et des mitogènes du rapport cycline D1/p27
Comment les tissus savent quand croître
Lorsque vous coupez la peau ou endommagez un organe, les cellules voisines quittent brièvement un état de repos pour passer en mode réparation, se multipliant juste assez pour colmater la blessure avant de s’arrêter à nouveau. Cet article pose une question apparemment simple : comment les cellules savent-elles quand commencer et, tout aussi important, quand arrêter de se diviser pour que la guérison se fasse sans dériver vers une prolifération du type cancéreux ? Les chercheurs mettent au jour un jeu d’équilibre intégré dans les cellules qui active un signal de croissance puissant uniquement de façon temporaire puis le désactive de manière fiable.
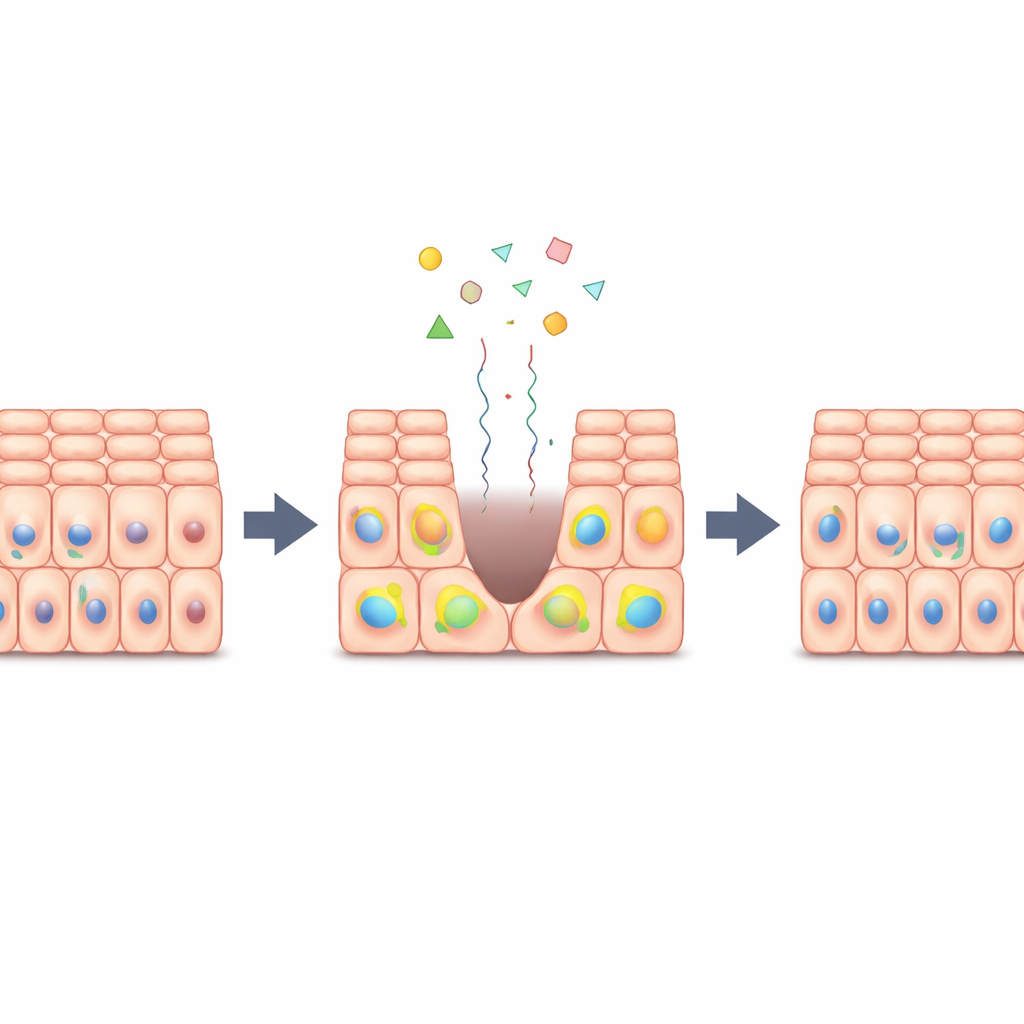
Un feu de circulation pour la division cellulaire
Dans de nombreux tissus, un système moléculaire appelé Hippo–YAP agit comme un contrôleur de la circulation pour la croissance cellulaire. Lorsque les tissus sont denses et intacts, Hippo maintient une protéine appelée YAP hors du noyau, et les cellules restent quiescentes. Lorsque le tissu est blessé ou étiré, Hippo se relâche, YAP migre dans le noyau et les cellules sont incitées à se diviser. Les auteurs ont étudié des milliers de cellules épithéliales individuelles cultivées en feuillets plats et les ont exposées à différentes combinaisons de facteurs de croissance (mitogènes), de médicaments et de densités cellulaires. Ils ont constaté que les signaux des facteurs de croissance, l’encombrement physique et l’activité de YAP ne fonctionnent pas isolément ; au contraire, ils convergent vers un point de décision interne unique qui détermine si chaque cellule réentrera dans le cycle cellulaire.
Le rapport clé à l’intérieur de la cellule
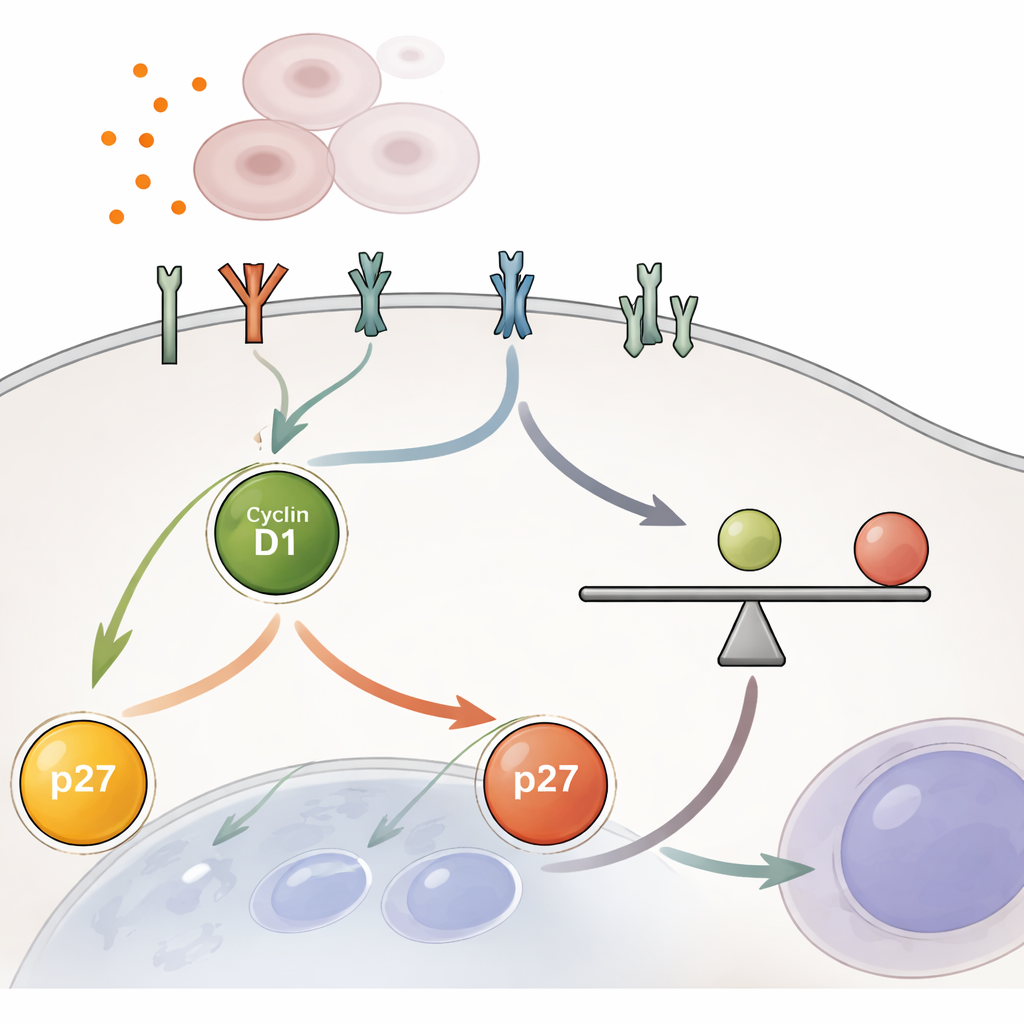
La découverte centrale est que les cellules décident de se diviser sur la base d’un rapport entre deux protéines dans le noyau pendant la phase précoce de croissance connue sous le nom de G1. Une protéine, la cycline D1, pousse la cellule vers la division ; l’autre, p27, fait office de frein. Plutôt que la quantité absolue de l’une ou l’autre, c’est le rapport cycline D1/p27 qui compte. Lorsque ce rapport dépasse un seuil critique, une protéine de garde appelée Rb devient phosphorylée et libère la machinerie nécessaire à la réplication de l’ADN, permettant à la cellule d’avancer. En dessous de ce seuil, la cellule reste à l’état de repos. L’équipe a montré qu’activer artificiellement YAP, ou bloquer les freins en amont de la voie Hippo, augmentait de façon constante le rapport cycline D1/p27 et poussait les cellules au-delà de ce seuil, même dans des conditions qui normalement les maintiennent silencieuses.
Comment les signaux externes sont amplifiés puis estompés
YAP n’agit pas seul : il augmente la réceptivité de la cellule aux signaux de croissance venant de l’extérieur. Les chercheurs ont utilisé le séquençage ARN et des mesures protéiques pour montrer que YAP accroît le nombre et l’activité des protéines réceptrices à la surface cellulaire, y compris des membres de la famille EGFR et d’autres récepteurs tyrosine kinases. Ces récepteurs alimentent des voies de croissance classiques telles que MEK–ERK et, dans une moindre mesure, mTOR, qui augmentent la cycline D1 et abaissent p27 pour orienter le rapport interne vers la division. Il est important de noter que cette amplification n’est pas un interrupteur marche/arrêt instantané. Même après blocage de l’activité de YAP, la signalisation réceptrice renforcée et le rapport cycline D1/p27 élevé décroissent lentement sur plusieurs heures, donnant aux cellules le temps d’achever une série limitée de divisions avant que les freins ne soient pleinement réappliqués.
Dispositifs de sécurité intégrés contre la croissance incontrôlée
L’étude explore également ce qui se passe lorsque les freins naturels sur YAP sont affaiblis, comme c’est le cas dans certaines tumeurs. L’élimination d’une protéine de jonction appelée Merlin, qui aide normalement à activer Hippo, a partiellement libéré YAP et rendu les cellules plus sensibles aux facteurs de croissance. Pourtant, ces cellules pouvaient encore être arrêtées par une densité cellulaire locale très élevée, qui rétablissait l’inhibition de contact et faisait baisser le rapport cycline D1/p27. Cela montre que le système comporte plusieurs couches de contrôle : les récepteurs de surface, Hippo–YAP et le rapport cycline D1/p27 contribuent tous à la décision de savoir si les cellules poursuivent leur division.
Pourquoi cela importe pour la cicatrisation et le cancer
Pour un lecteur général, le message essentiel est que la réparation tissulaire est portée par un programme de croissance transitoire et auto-limitant. L’activation de YAP et l’augmentation de la signalisation réceptrice font monter le rapport cycline D1/p27 au-dessus d’un seuil afin que les cellules puissent se diviser et réparer les lésions. À mesure que les cellules se divisent et que le tissu redevient dense, l’inhibition de contact éteint progressivement YAP et la signalisation des récepteurs, le rapport chute et la division s’arrête. Lorsque ce mécanisme temporel est rompu — par exemple par une activation persistante de YAP ou la perte de freins en amont — les cellules peuvent devenir insensibles à l’encombrement et continuer à se diviser, un pas vers le cancer. Comprendre ce système décisionnel fondé sur un rapport pourrait aider à concevoir des thérapies régénératives qui stimulent la réparation en toute sécurité sans déclencher une croissance incontrôlée.
Citation: Ferrick, K.R., Upadhya, S.W., Fan, Y. et al. Transient proliferation by reversible YAP and mitogen control of the cyclin D1/p27 ratio. Commun Biol 9, 340 (2026). https://doi.org/10.1038/s42003-026-09590-2
Mots-clés: signalisation YAP, contrôle du cycle cellulaire, régénération tissulaire, inhibition de contact, équilibre cycline D1 p27