Clear Sky Science · fr
Signatures neuronales distinctes de la dynamique des populations hippocampiques lors d’une locomotion sur place
Comment le cerveau suit le mouvement sans aller nulle part
Même lorsque vous courez sur un tapis roulant sans avancer, votre cerveau parvient malgré tout à suivre la vitesse, la distance parcourue et la durée du mouvement. Cette étude examine comment une région clé du cerveau, l’hippocampe — surtout connue pour la mémoire et la navigation — traite différents types de mouvement, depuis la course soutenue provoquée par un stimulus jusqu’aux mouvements agités et presque immobiles. Comprendre ces « codes » internes du mouvement peut éclairer la façon dont le cerveau construit notre sens de l’espace, du temps et de l’action, et comment ces mécanismes peuvent se détériorer avec l’âge ou dans la maladie.
Un examen attentif du cerveau en mouvement
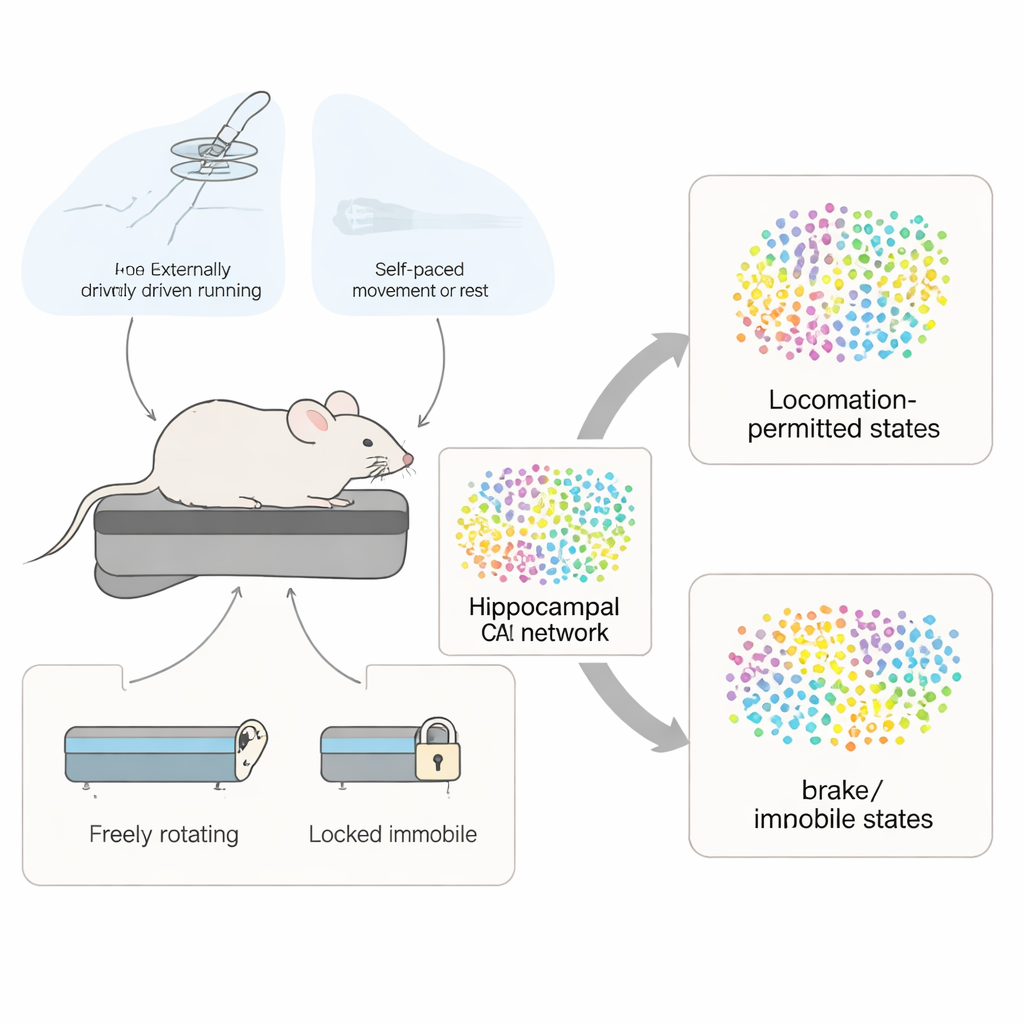
Pour observer de nombreuses cellules cérébrales tout en gardant la situation strictement contrôlée, les chercheurs ont travaillé avec des souris dont la tête était légèrement fixée au-dessus d’un tapis roulant simple et non motorisé. Un jet d’air léger dans le dos poussait les animaux à courir ; couper le flux d’air leur permettait de ralentir ou de s’arrêter d’eux-mêmes. Lors de certaines sessions, la bande pouvait tourner librement, autorisant des courses à plein appui sur place. Lors d’autres, un frein bloquait la bande de sorte que seuls de légers mouvements de pattes étaient possibles. En continu, un microscope détectant des éclairs de calcium dans les cellules nerveuses a enregistré l’activité de centaines de neurones hippocampiques, permettant à l’équipe d’inférer quand chaque cellule devenait plus ou moins active.
Différents types de course, différents rôles neuronaux
Sur le plan comportemental, le jet d’air a généré deux états de mouvement bien distincts. Pendant les périodes avec air sur une bande libre, les souris atteignaient rapidement et maintenaient des vitesses relativement élevées, se comportant comme une personne maintenant le rythme sur un tapis roulant. Quand l’air était coupé, elles continuaient de bouger un moment puis passaient à des épisodes plus lents, irréguliers et auto-initiés. Sur une bande verrouillée, les mêmes rafales d’air produisaient seulement de petits mouvements sur place, mais ceux-ci différaient encore entre phases d’air allumé et éteint. Les chercheurs se sont demandé dans quelle mesure l’activité de chaque cellule hippocampique était liée à trois quantités simples : le temps écoulé, la distance parcourue (ou, avec le frein, l’amplitude du mouvement sur place) et la vitesse du déplacement.
Codes nets et simples après le stimulus
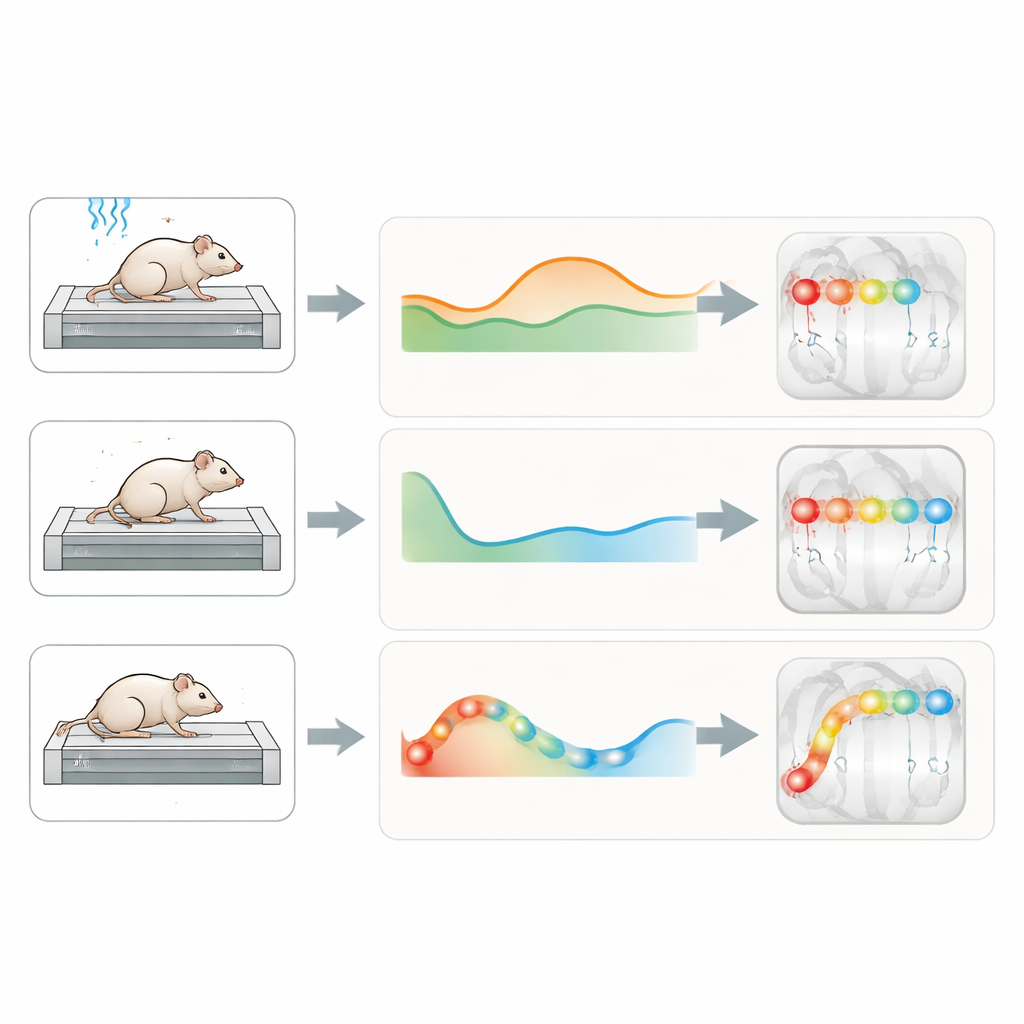
Dans toutes les conditions, davantage de cellules étaient actives et clairement liées aux variables de mouvement pendant les périodes post‑stimulus avec l’air coupé, lorsque les animaux se mouvaient de leur propre initiative. En contrôlant le fait que les phases sans air duraient simplement plus longtemps, l’équipe a trouvé que la course sous air-on recrutait en réalité un sous-ensemble de cellules plus fiable — mais sur la fenêtre plus longue de l’air-off, de nombreux neurones supplémentaires entraient en jeu. Au sein de cette population active, la plupart des cellules se sont révélées « spécialistes » : leur activité était liée principalement à une seule caractéristique — temps, distance ou vitesse — plutôt qu’à un mélange complexe des trois. Cette propension à un réglage simple, centré sur une seule variable, était la plus marquée pendant l’air-off, ce qui suggère qu’une fois le stimulus moteur terminé, les réseaux hippocampiques passent dans un mode qui met en avant des aspects spécifiques du mouvement en cours.
La vitesse précède, le temps et la distance suivent
En examinant le timing de l’activité, un schéma frappant est apparu. Les cellules dont l’activité reflétait la vitesse atteignaient généralement leur pic d’activation plus tôt après le début ou la fin du flux d’air que les cellules qui suivaient le temps ou la distance. Autrement dit, les signaux liés à la vitesse s’allumaient rapidement autour de l’événement sensoriel qui lançait ou arrêtait la course, tandis que les codes de temps et de distance se construisaient plus tard, à mesure que le mouvement se déroulait. En immobilité forcée, les cellules étaient encore majoritairement des spécialistes, désormais réglées soit sur le temps soit sur de subtils mouvements sur place, les signaux de mouvement sur place étant particulièrement marqués après l’arrêt de l’air. Cela suggère un rôle de l’hippocampe dans la surveillance même de tentatives de mouvement infimes lorsque le déplacement en avant est empêché.
Motifs de groupe stables malgré des individus changeants
Au niveau des cellules individuelles, la distribution de ce que chaque neurone codait était étonnamment fluide : une cellule qui suivait la vitesse dans une configuration pouvait coder le temps, la distance ou rien du tout dans une autre. Pourtant, quand les auteurs ont regardé la population dans son ensemble, ils ont trouvé une structure ordonnée. Les groupes de cellules actifs durant la même phase — air-on ou air-off — se ressemblaient davantage entre eux que des groupes entre phases, et des motifs formaient des grappes distinctes pour les conditions de course libre versus freinée. Cela suggère que l’hippocampe conserve une « armature » stable d’organisation de population tout en réaffectant de façon flexible les rôles des neurones individuels selon le contexte de mouvement.
Ce que cela signifie pour notre perception interne du mouvement
Synthétiquement, l’étude montre que l’hippocampe ne s’appuie pas sur un ensemble fixe de cellules pour suivre le mouvement. Il rééquilibre dynamiquement des signaux simples portant sur la vitesse, le temps, la distance et même de petits mouvements sur place selon que le mouvement est conduit par un stimulus externe ou auto‑initié, et selon que le corps est libre de bouger ou maintenu immobile. Les signaux de vitesse apparaissent en premier autour des événements sensoriels importants, tandis que des codes temporels et de distance plus précis émergent au fur et à mesure du comportement. Malgré cette rotation au niveau des cellules individuelles, le schéma global d’activité reste bien organisé et lié à l’état comportemental. Un tel système à la fois flexible et structuré pourrait sous-tendre notre capacité à former des souvenirs qui lient où nous étions, comment nous nous sommes déplacés et quand les événements se sont produits — même lorsque nous n’avons en réalité pas bougé de place.
Citation: Inayat, S., McAllister, B.B., Whishaw, I.Q. et al. Distinct neural signatures of hippocampal population dynamics during locomotion-in-place. Sci Rep 16, 10372 (2026). https://doi.org/10.1038/s41598-026-41049-6
Mots-clés: hippocampe, locomotion, codage neuronal, dynamique de population, intégration sensorimotrice