Clear Sky Science · fr
Signature dynamique du compromis activité-stabilité dans l'évolution des lactamases
Pourquoi c’est important pour la résistance aux antibiotiques
La résistance aux antibiotiques paraît souvent comme une boîte noire : les bactéries « changent d’une façon ou d’une autre » et les médicaments ne fonctionnent plus. Cette étude ouvre cette boîte pour l’une des enzymes de résistance les plus connues au monde, la bêta‑lactamase TEM‑1, qui permet aux bactéries de détruire les antibiotiques de type pénicilline. En observant comment la forme et les mouvements de cette enzyme évoluent pour cibler des médicaments plus récents, les auteurs expliquent pourquoi des mutations qui augmentent l’activité affaiblissent souvent la stabilité, et comment l’évolution trouve des moyens ingénieux d’équilibrer les deux.
D’un destructeur de pénicilline à un briseur de médicaments plus large
TEM‑1 est à l’origine excellente pour dégrader les anciennes pénicillines mais peu efficace contre des antibiotiques plus récents et encombrants comme le céfotaxime. Dans de nombreuses souches cliniques, une mutation clé unique appelée G238S apparaît près du site actif de l’enzyme, la poche où les antibiotiques sont clivés. Cette mutation améliore de façon spectaculaire la capacité de l’enzyme à détruire le céfotaxime, tout en n’affectant que modestement son rôle original sur la pénicilline. Les auteurs montrent que G238S n’élargit pas simplement la poche ; elle réorganise la façon dont plusieurs boucles et hélices environnantes bougent, créant une nouvelle conformation fonctionnelle qui accueille mieux les molécules volumineuses.
Le mouvement des protéines comme bouton de réglage évolutif
Grâce à des techniques avancées de résonance magnétique nucléaire (RMN), les chercheurs ont mesuré comment différentes parties de TEM‑1 bougent sur des échelles de temps allant de la trillionième à la millième de seconde. Le TEM‑1 sauvage est assez rigide, ce qui l’aide à traiter efficacement ses substrats d’origine. G238S préserve la plupart de cette rigidité rapide mais introduit des mouvements lents et finement réglés dans plusieurs parois du site actif. Ces mouvements sont assez rapides pour suivre la chimie de l’enzyme sans devenir si désordonnés qu’ils perturbent les résidus catalytiques critiques. Le résultat est une « fenêtre optimisée » de flexibilité : juste assez de mouvement pour ouvrir la poche au céfotaxime, tout en maintenant l’alignement correct de la machinerie chimique centrale.
Balancer deux formes au lieu d’en choisir une
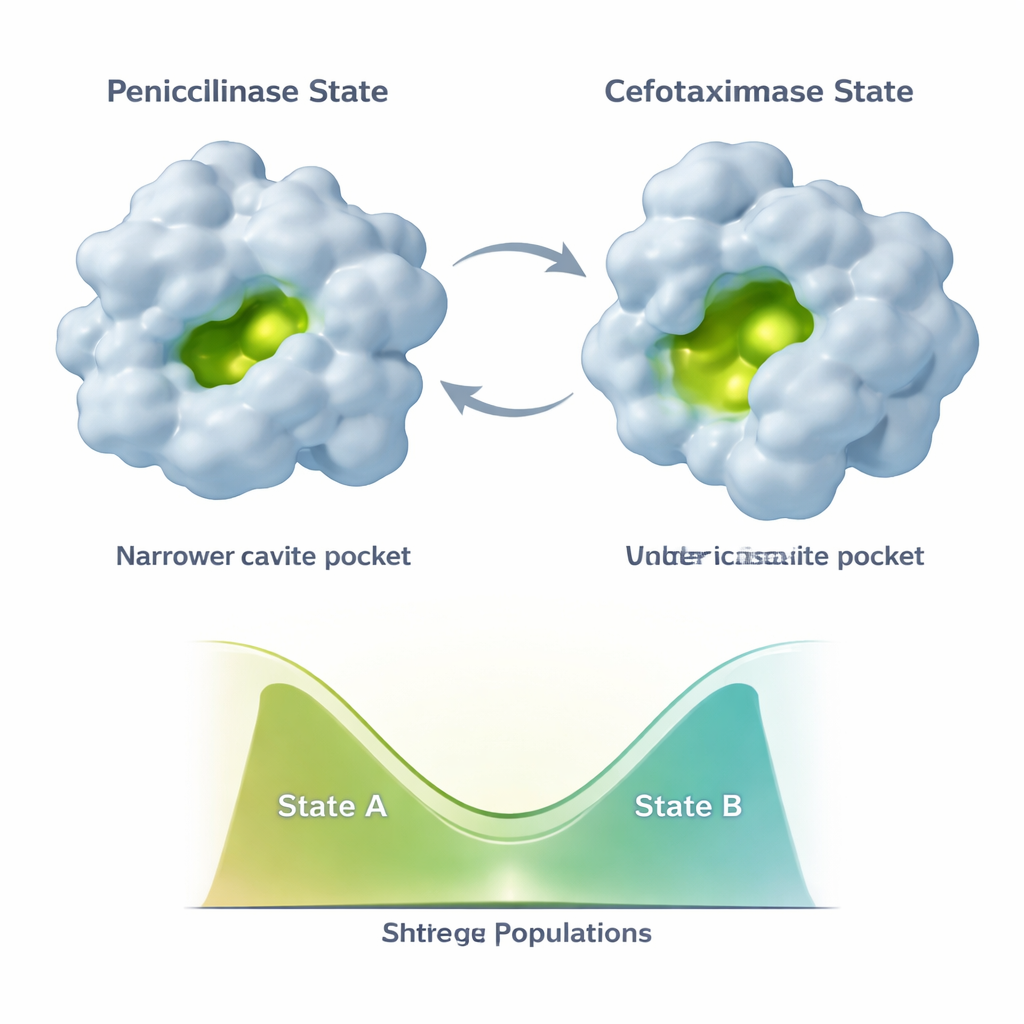
L’évolution ne fige pas TEM‑1 dans une seule nouvelle forme. Au contraire, l’enzyme échantillonne au moins deux principales conformations : un état « pénicillinase » ressemblant à la structure originale et un état plus ouvert « céfotaximase » mieux adapté aux médicaments récents. Des mutations additionnelles qui apparaissent ensuite, comme E104K et A42G, ont un effet subtil. Plutôt que de rendre simplement la nouvelle forme favorable plus stable, elles rééquilibrent le mélange des deux états. Les données RMN montrent que différentes parties du site actif et de l’ossature porteuse peuvent faire varier leurs populations indépendamment le long de ce continuum à deux états. Cela conduit à un ensemble combinatoire de variantes enzymatiques, chacune avec un mélange différent de conformations de type pénicilline et de type céfotaxime, et par conséquent des profils catalytiques distincts.
Points faibles cachés et réparations à distance
Les mutations qui améliorent l’activité entraînent souvent un coût caché : elles rendent la protéine moins stable. Plutôt que de s’intéresser uniquement au dépliement complet, l’équipe a cartographié la stabilité locale au niveau de courts segments en utilisant l’échange hydrogène–déutérium couplé à la spectrométrie de masse. G238S destabilise non seulement des boucles proches mais aussi des hélices et régions en feuillet éloignées qui forment une ossature structurelle. Certaines de ces régions se recoupent avec une poche allostérique « cryptique »—une ouverture rarement visitée dans le cœur de la protéine qui peut lier de petites molécules et atténuer l’activité. G238S facilite l’ouverture de cette poche, incorporant en quelque sorte une légère fonction d’auto‑inhibition dans l’enzyme. Les mutations ultérieures, en particulier A42G, renforcent ce réseau d’ossature affaibli, améliorant la stabilité locale autour de trois hélices en interaction sans supprimer les dynamiques bénéfiques du site actif. Autrement dit, l’évolution colmate les points faibles structurels à distance au lieu d’annuler l’innovation initiale.
Ce que l’étude révèle sur la stratégie de l’évolution
Pour un lecteur non spécialiste, le message central est que des protéines comme TEM‑1 n’acquièrent pas la résistance en actionnant un simple interrupteur marche/arrêt. Chaque mutation modifie légèrement la façon dont l’enzyme respire, fléchit et répartit son temps entre différentes poses de travail. G238S ouvre la porte à une nouvelle fonction—dégrader des antibiotiques plus récents—mais crée aussi des fragilités locales et un état partiellement auto‑inhibiteur. Les mutations secondaires agissent comme des renforts prudents, stabilisant l’ossature et ajustant finement l’équilibre entre anciennes et nouvelles conformations pour que l’enzyme reste à la fois active et durable. Cette vision dynamique de l’évolution, où les mouvements et les faiblesses locales comptent autant que la structure statique, peut aider à guider la conception de futurs antibiotiques et de médicaments ciblant les enzymes, plus difficiles à contourner pour les bactéries.
Citation: Arcia, E., Keramisanou, D., Jacobs, L.M.C. et al. Dynamic signature of activity-stability tradeoff in lactamase evolution. Nat Commun 17, 1884 (2026). https://doi.org/10.1038/s41467-026-68620-z
Mots-clés: résistance aux antibiotiques, bêta-lactamase, évolution des protéines, dynamique enzymatique, stabilité des protéines