Clear Sky Science · fr
Rôles distincts des sous-types de la couche 5 corticale dans l’apprentissage associatif
Comment le cerveau apprend qu’un toucher annonce une récompense
Imaginez apprendre qu’une légère tape sur votre poignet signifie que le dessert arrive. Votre cerveau doit lier un simple toucher à une récompense future. Cette étude explore l’intérieur du cerveau de la souris pour voir comment deux types différents de cellules nerveuses coopèrent pour former ces associations, révélant comment l’apprentissage quotidien se répartit entre « ce qui s’est passé » et « ce que cela veut dire pour moi ». 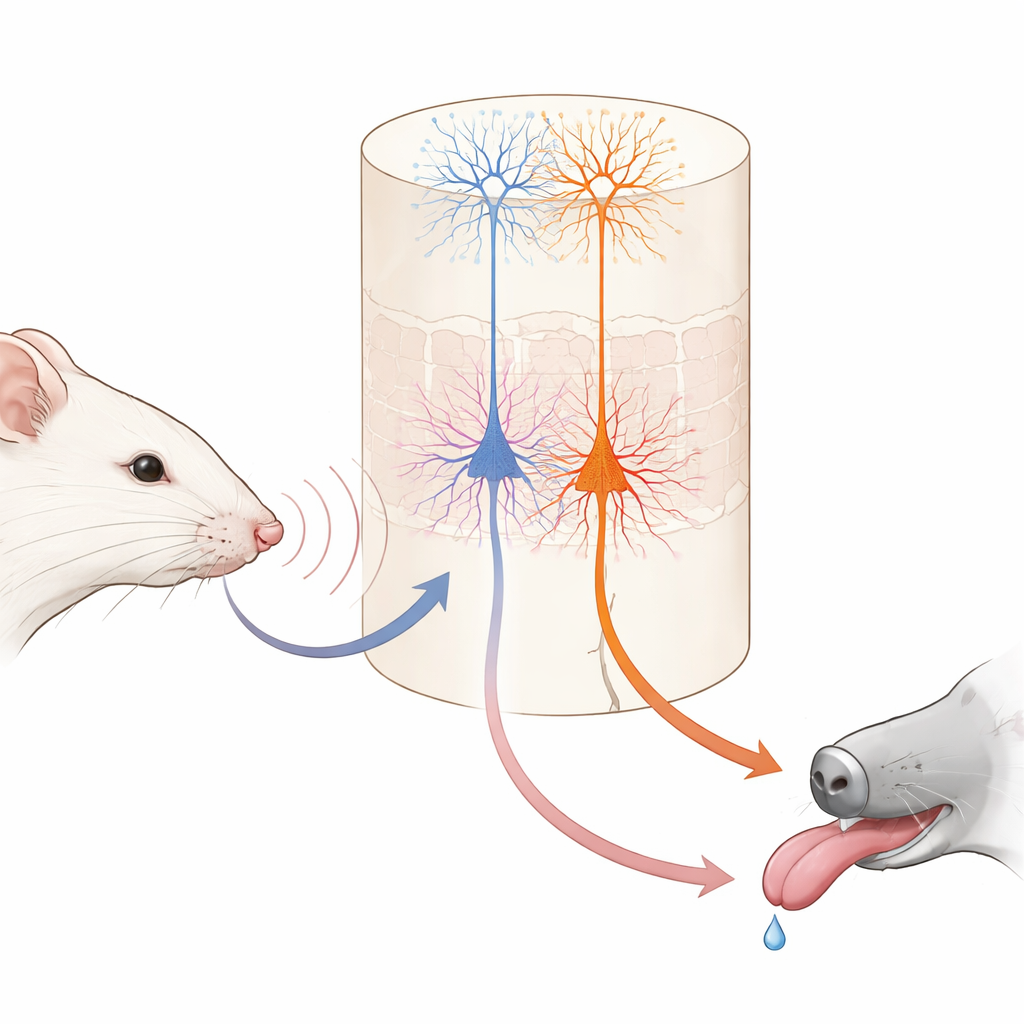
Un jeu simple de vibrisses pour étudier l’apprentissage
Les chercheurs ont entraîné des souris tête-fixées à un jeu simple utilisant leurs vibrisses faciales. À chaque essai, une seule vibrisse était vibrée à l’une des deux vitesses. Une vibration était suivie, après un bref délai, d’une petite goutte d’eau ; l’autre n’était jamais récompensée. Au début, les souris léchaient en anticipation lors des deux types d’essais, ne sachant pas encore quelle sensation sur la vibrisse signalait l’eau. En quelques jours, elles ont progressivement appris à lécher tôt seulement lorsque la vibration « bonne » se produisait, et à se retenir quand la vibration non récompensée apparaissait. Lorsque les scientifiques ont temporairement désactivé la zone primaire du toucher du cerveau pendant l’entraînement, cet apprentissage a en grande partie disparu, montrant que cette région sensorielle est importante pour construire l’association, même si des souris expertes pouvaient plus tard exécuter la tâche sans elle.
Deux types de cellules de sortie avec des tâches très différentes
Dans cette zone tactile, une couche profonde connue sous le nom de couche 5 contient deux types principaux de cellules nerveuses de sortie. Un groupe, appelé ici cellules IT, envoie des signaux vers d’autres régions corticales des deux hémisphères cérébraux. L’autre groupe, les cellules ET, envoie principalement des signaux vers les structures sous-corticales impliquées dans l’action et la récompense. En utilisant des souris génétiquement modifiées et une imagerie biphotonique haute résolution, les auteurs ont pu surveiller sélectivement l’activité de chaque type cellulaire à travers les extrémités de leurs longs branches en forme d’arbre. Avant l’apprentissage, les cellules IT répondaient déjà fortement et de manière fiable aux vibrations des vibrisses, et leur activité combinée pouvait distinguer précisément les deux vitesses de vibration. Les cellules ET, en revanche, répondaient de manière plus faible et moins cohérente aux stimuli, n’offrant qu’une lecture floue de la vibration survenue.
Sensations stables versus attentes croissantes
À mesure que les souris apprenaient, les cellules IT se comportaient comme des observateurs fiables. Leurs réponses restaient étroitement liées au moment du déplacement de la vibrisse et changeaient peu d’un jour à l’autre. Elles continuaient à encoder quelle vibration avait eu lieu, indépendamment de son caractère prédictif d’une récompense. Les cellules ET, toutefois, transformèrent leur comportement. Au lieu de simplement décharger au début du stimulus, leur activité augmentait progressivement pendant la vibration et le court délai, culminant autour du moment attendu de la délivrance d’eau. Cette montée s’amplifiait parallèlement aux léchages anticipatoires des animaux et devint meilleure pour prédire si un essai se terminerait par un léchage que pour rendre compte précisément du stimulus. Des cellules ET individuelles entraient et sortaient du groupe actif au fil des jours, mais au niveau de la population le schéma devenait plus cohérent, suggérant un code flexible mais convergent pour l’attente de la récompense. 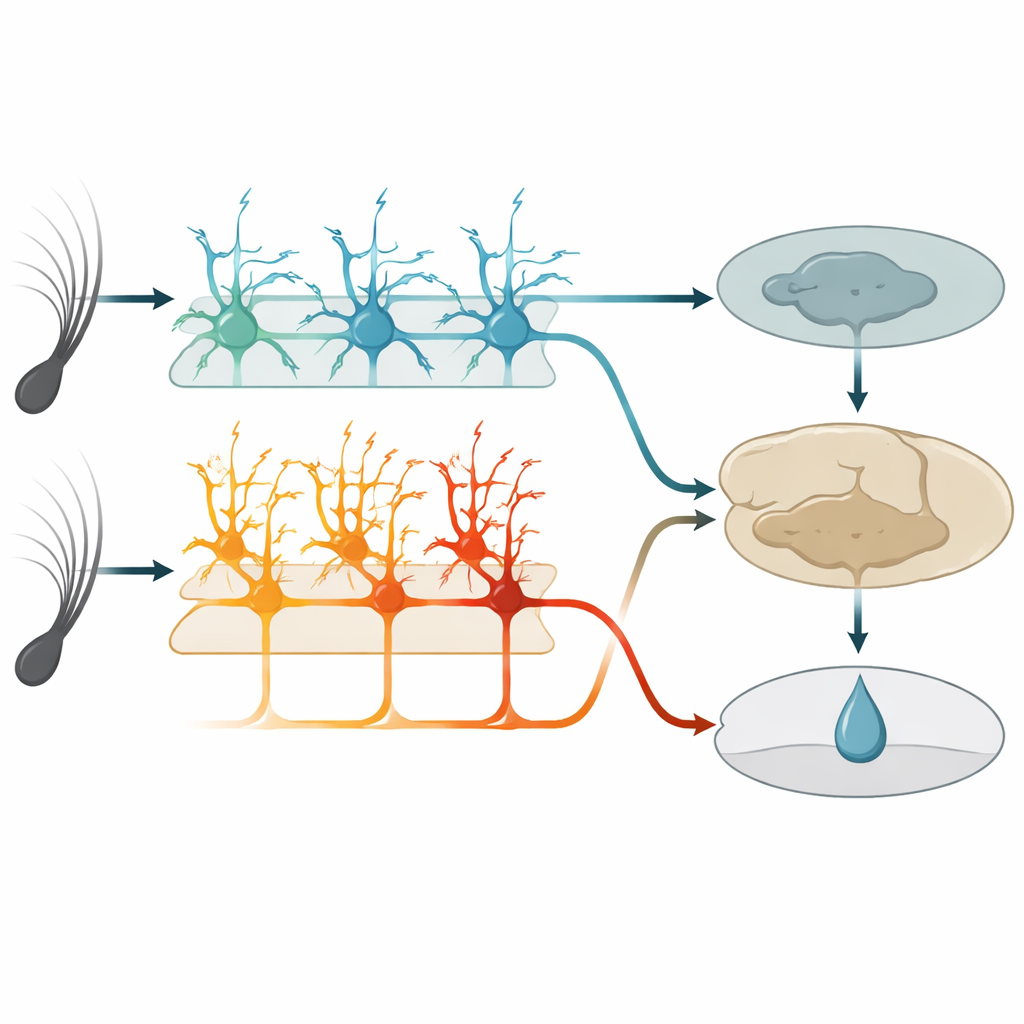
Désactiver chaque type cellulaire révèle une division du travail
Pour tester la fonction, l’équipe a utilisé des outils chimio-génétiques pour atténuer sélectivement soit les cellules IT soit les cellules ET pendant l’entraînement. Lorsque les cellules IT furent silencées, les souris présentaient globalement moins de léchages anticipatoires et ne parvenaient pas à établir une différence nette entre les vibrations récompensées et non récompensées. Lorsque les cellules ET furent silencées, les souris eurent l’effet inverse : elles léchaient excessivement pour les deux vibrations, en particulier pour celle non récompensée, et avaient du mal à affiner leur comportement même si elles léchaient toujours avec vigueur. La mise hors fonction de l’un ou l’autre groupe après maîtrise de la tâche n’altérait plus les performances, ce qui implique qu’une fois que d’autres régions cérébrales ont stocké l’association, cette aire sensorielle et ses sorties de la couche 5 deviennent moins critiques pour l’exécution de la réponse apprise.
Un modèle d’apprentissage qui reflète le comportement cérébral
Les auteurs ont construit un modèle informatique de type apprentissage par renforcement pour interpréter ces résultats. Dans le modèle, un réseau de type IT fournit des représentations sensorielles stables qui aident à estimer la « valeur » de chaque stimulus—la probabilité qu’il soit suivi d’une récompense. Une voie de type ET relaie cette valeur prédite vers un circuit en aval qui la compare avec la récompense réelle, générant une erreur de prédiction qui ajuste les estimations de valeur futures. Le blocage des voies IT ou ET dans le modèle reproduisait les échecs d’apprentissage distincts observés chez les souris : sans entrée de type IT, l’apprentissage était lent et faible pour les deux stimuli ; sans sortie de type ET, un apprentissage initial se produisait mais le système n’arrivait pas à réduire correctement les réponses au signal non récompensé. Le modèle rendait aussi compte du fait que, avec le temps, des régions non sensorielles puissent prendre le relais de la performance, en accord avec les expériences.
Ce que cela signifie pour l’apprentissage quotidien
En termes simples, cette étude suggère que lorsque nous apprenons qu’un certain visage, son ou toucher prédit quelque chose de bon ou de mauvais, différents ensembles de neurones corticaux profonds partagent le travail. Un ensemble garde une trace fidèle de « ce qui s’est passé » dans le monde, tandis qu’un autre en vient progressivement à signaler « ce que j’attends qui arrive ensuite » et aide à affiner le comportement en comparant ces attentes à la réalité. Ensemble, ils forment un pont entre la sensation brute et des actions flexibles fondées sur l’expérience, offrant une image plus claire de la façon dont le cerveau soutient les habitudes, l’apprentissage par récompense, et possiblement comment ces processus se dérèglent dans la maladie.
Citation: Moberg, S., Garibbo, M., Mazo, C. et al. Distinct roles of cortical layer 5 subtypes in associative learning. Nat Commun 17, 2648 (2026). https://doi.org/10.1038/s41467-026-68307-5
Mots-clés: apprentissage associatif, cortex somatosensoriel, neurones de la couche 5, attente de récompense, apprentissage par renforcement