Clear Sky Science · es
La vía perforante y los aferentes CA3‑colaterales de Schaffer coordinan la regulación del aprendizaje espacial
Por qué es importante orientarse
Tanto si se trata de un ratón buscando una plataforma oculta como de una persona que se desplaza por una ciudad desconocida, el cerebro convierte continuamente vistas, sonidos y el propio movimiento en un mapa interno. Este estudio investiga cómo dos regiones cerebrales clave, la corteza entorrinal y el hipocampo, trabajan en conjunto para sostener esta habilidad. Observando la actividad en fibras nerviosas diminutas en tiempo real y estimulando o silenciando con precisión vías específicas en ratones, los investigadores revelan cómo estas áreas se coordinan para formar y estabilizar memorias espaciales—con hallazgos que podrían, con el tiempo, ayudar a entender y tratar trastornos de la memoria.
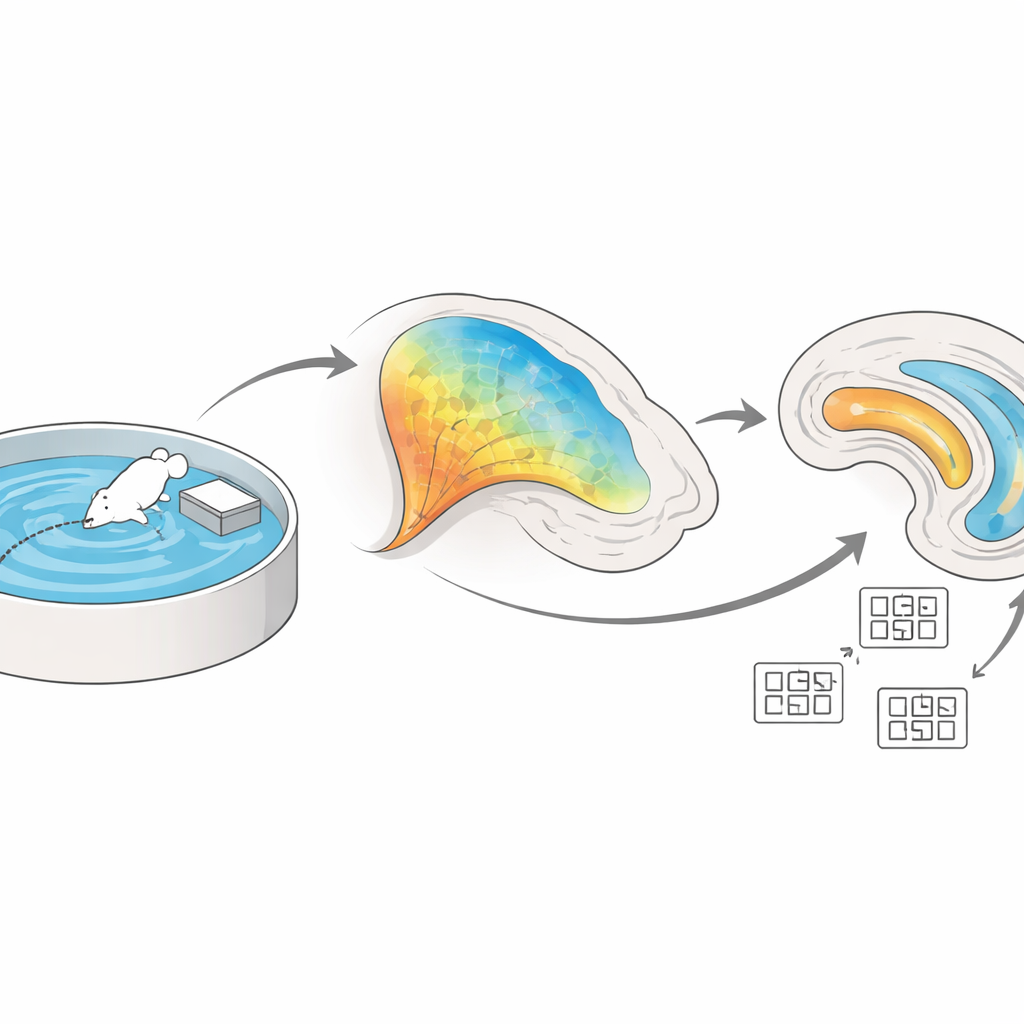
Dos centros clave del GPS interno del cerebro
El hipocampo y la vecina corteza entorrinal se sitúan en lo profundo del lóbulo temporal y constituyen el núcleo de nuestro sistema de navegación. Dentro del hipocampo, un circuito interno enlaza una zona llamada CA3 con otra llamada CA1 a través de fibras conocidas como colaterales de Schaffer. Al mismo tiempo, la corteza entorrinal envía una corriente separada de información directamente a CA1 mediante la llamada vía perforante. Los autores se propusieron determinar cómo cooperan estas dos entradas cuando un animal aprende la distribución de un espacio, y cómo los cambios en sus conexiones—conocidos en general como “plasticidad”—apoyan la construcción de un mapa espacial estable.
Observar el aprendizaje fibra por fibra
Para seguir este proceso en animales que se comportan libremente, el equipo empleó fotometría de fibra, una técnica que informa sobre la actividad nerviosa mediante destellos de luz fluorescente. Diseñaron ratones de modo que las neuronas CA3 que proyectan a CA1 brillaran al activarse, y luego entrenaron a los animales en el laberinto acuático de Morris, una prueba clásica en la que el ratón debe aprender la ubicación oculta de una pequeña plataforma en una piscina. Durante las primeras pruebas, las fibras CA3–CA1 mostraron una fuerte actividad mientras los animales buscaban; a medida que los ratones se hicieron diestros y encontraron la plataforma más rápido, esa actividad disminuyó gradualmente. El patrón sugiere que estas conexiones se implican especialmente mientras el cerebro codifica por primera vez la disposición del entorno, y luego se asientan en una representación más eficiente y estable una vez que la memoria se ha establecido.
Cómo la entrada upstream potencia o atenúa el aprendizaje
Los científicos preguntaron a continuación cómo las señales de la corteza entorrinal influyen en este circuito hipocampal. Usando proteínas sensibles a la luz para activar neuronas entorrinales mientras registraban desde las fibras CA3–CA1, demostraron que estimular la entrada desde la corteza entorrinal medial aumentaba de forma fiable la actividad en esas conexiones hipocampales. Por el contrario, cuando atenúan químicamente las señales entorrinales que llegan a CA1 durante el entrenamiento en el laberinto acuático, la actividad CA3–CA1 se debilitó y los ratones aprendieron la ubicación de la plataforma más despacio y con menos precisión. En un conjunto complementario de experimentos, los autores registraron directamente desde neuronas entorrinales que proyectan a CA1 y observaron que su actividad aumentaba a lo largo de los días de entrenamiento, reflejando la mejora del rendimiento de los animales. En conjunto, estos resultados indican que señales entorrinales fuertes y bien sincronizadas son necesarias para ajustar correctamente los circuitos hipocampales para la navegación.
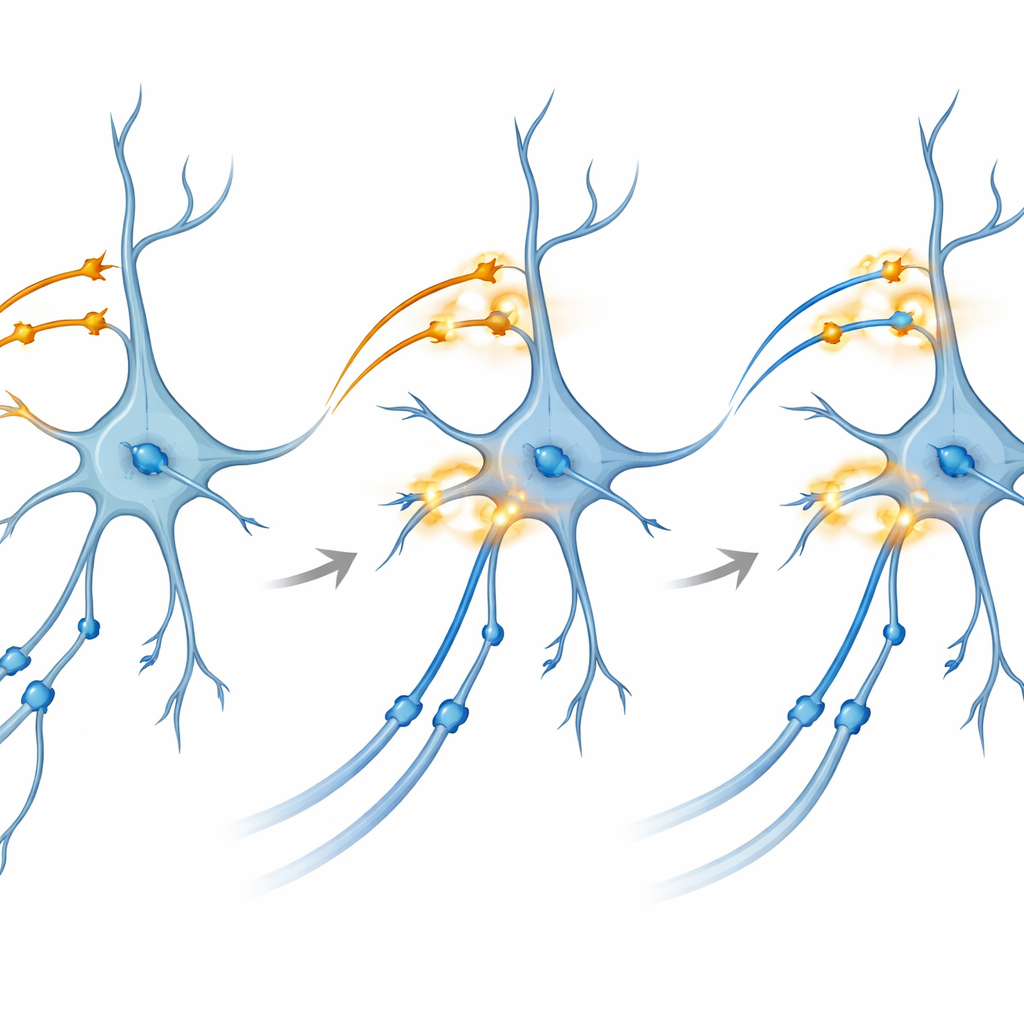
Acercándose a cómo se fortalecen las conexiones
Para sondear el mecanismo subyacente, el equipo recurrió a cortes de cerebro mantenidos vivos en una placa. Allí pudieron controlar por separado proteínas activadas por luz en CA3 y en las entradas entorrinales a CA1. Sorprendentemente, la estimulación con ráfagas en ritmo theta—patrones que imitan el disparo rítmico natural—aplicada a cualquiera de las vías por sí sola no produjo de forma fiable potenciación a largo plazo, el fortalecimiento duradero de las sinapsis que se considera base de la memoria. Sin embargo, cuando aplicaron una ráfaga con doble color cuidadosamente sincronizada que coactivaba ambas vías sobre CA1 al mismo tiempo, las conexiones CA3–CA1 mostraron una mejora robusta y persistente. Bloquear los receptores NMDA o canales de calcio específicos, dos puertas moleculares bien conocidas para la plasticidad, impidió este efecto, vinculando el fenómeno a rutas bioquímicas clásicas de la formación de memoria.
Qué significa esto para la memoria y la enfermedad
En conjunto, el estudio retrata el aprendizaje espacial como una danza coordinada entre las señales entrantes de la corteza entorrinal y el cableado interno del hipocampo. Las entradas entorrinales no solo transmiten información sobre la posición y el contexto; también actúan como un potente control que puede amplificar o atenuar la actividad en las fibras CA3–CA1 y, cuando se coactivan, inducir el fortalecimiento a largo plazo de esas sinapsis. Para un lector general, la conclusión clave es que el “GPS” del cerebro no depende de una sola zona, sino de una asociación en la que una región pone la escena y la otra fija los detalles. Las alteraciones de esta asociación—ya sea por envejecimiento, lesiones o enfermedades neurodegenerativas—podrían ayudar a explicar por qué las personas se desorientan en lugares que conocen, y dirigir estas vías podría ofrecer nuevas vías para preservar o restaurar la memoria espacial.
Cita: Huang, F., Temitayo Bello, S., Lau, S.H. et al. The perforant pathway and CA3-Schaffer collateral afferents coordinate to regulate spatial learning. Commun Biol 9, 364 (2026). https://doi.org/10.1038/s42003-026-09577-z
Palabras clave: memoria espacial, hipocampo, corteza entorrinal, plasticidad sináptica, navegación