Clear Sky Science · es
Sororin bloquea la puerta de salida del ADN del complejo cohesina para preservar la cohesión de las cromátidas hermanas
Mantener juntas de forma segura las cromosomas copiadas
Cada vez que una célula se divide, debe copiar su ADN y luego entregar un juego completo a cada célula hija. Si esas copias se separan demasiado pronto, el resultado puede ser cromosomas rotos, caos genético y enfermedad, incluido el cáncer. Este estudio revela cómo una pequeña porción proteica llamada cola de Sororin actúa como una cerradura física sobre una abrazadera con forma de anillo que mantiene unidas las copias hermanas del ADN, explicando cómo las células conservan los cromosomas emparejados hasta el momento exacto de la separación.
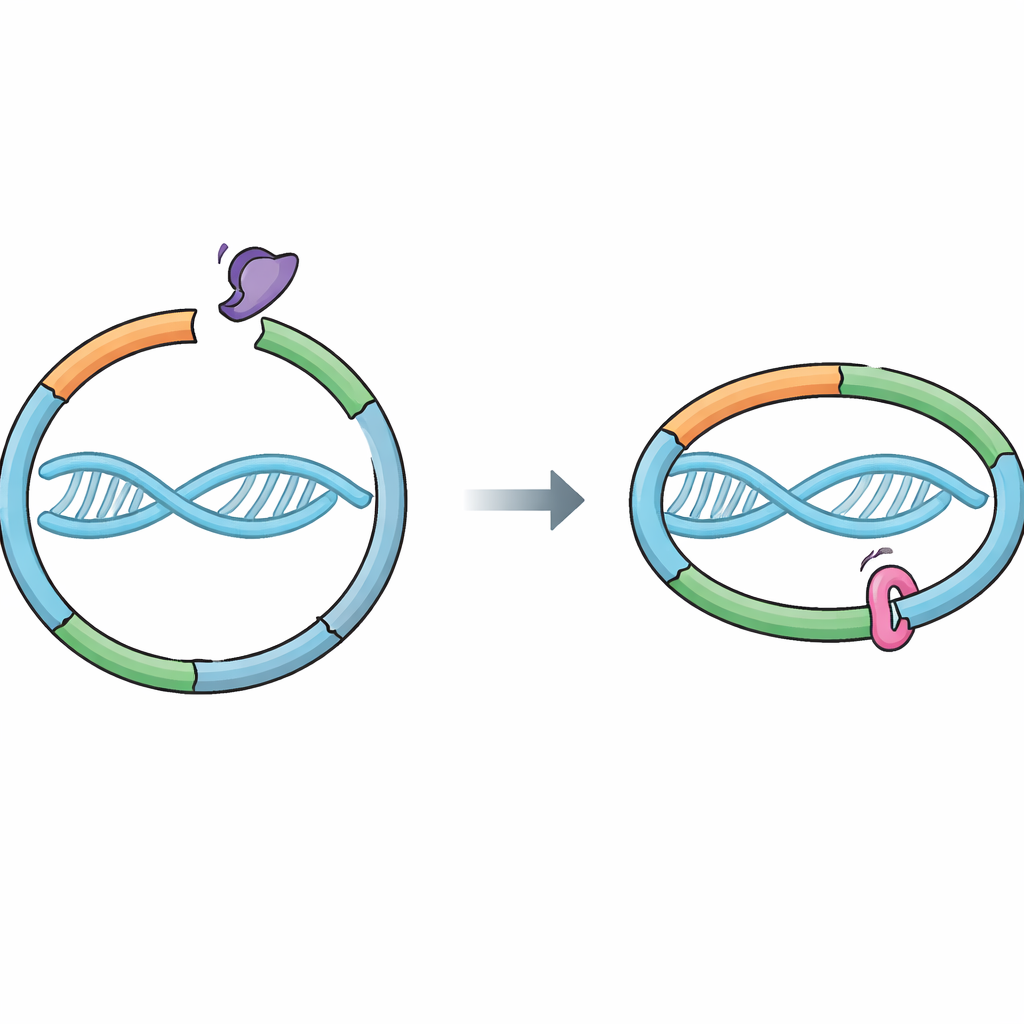
Un anillo molecular que protege nuestros genes
Las células dependen de una máquina proteica llamada cohesina para mantener juntas las cromátidas recién copiadas. La cohesina forma un anillo que rodea el ADN, uniendo las dos hebras hermanas creadas durante la replicación. Este enlace debe ser lo bastante fuerte para soportar los vaivenes de la división celular, pero liberable en el momento preciso para que las hermanas puedan separarse de forma ordenada. Una proteína asistente, Wapl, favorece la apertura de una “puerta de salida” específica en el anillo donde se encuentran dos partes de la cohesina, RAD21 y SMC3. La apertura de esta puerta permite que el ADN se deslice fuera y la cohesina se desprenda. Hasta ahora se pensaba que Sororin preservaba la cohesión principalmente bloqueando a la pareja de Wapl, Pds5, pero no quedaba claro si Sororin también tocaba directamente el anillo de cohesina.
Una cola diminuta con gran poder de sujeción
Los autores se centraron en el extremo extremo de la cola de Sororin: los últimos 30 aminoácidos, llamados región C‑terminal (CTR). Al recortar Sororin en fragmentos y probarlos en células humanas, hallaron que esta diminuta CTR es necesaria y suficiente para mantener emparejadas las cromátidas hermanas. Cuando la CTR se ancló de forma artificial en los centrómeros, los sitios centrales donde las hermanas permanecen unidas por más tiempo, casi rescató por completo la cohesión en células que carecían de Sororin completo. Por el contrario, cuando la CTR se unió a lo largo de la cromatina, la cohesina ya no pudo retirarse de manera eficiente, y los cromosomas no se compactaron ni se desenredaron correctamente. Estas células mostraron cromosomas mitóticos largos y difusos, puentes de ADN entre hermanas en separación y errores de segregación: problemas que se asemejan notablemente a los que ocurren cuando se elimina Wapl.
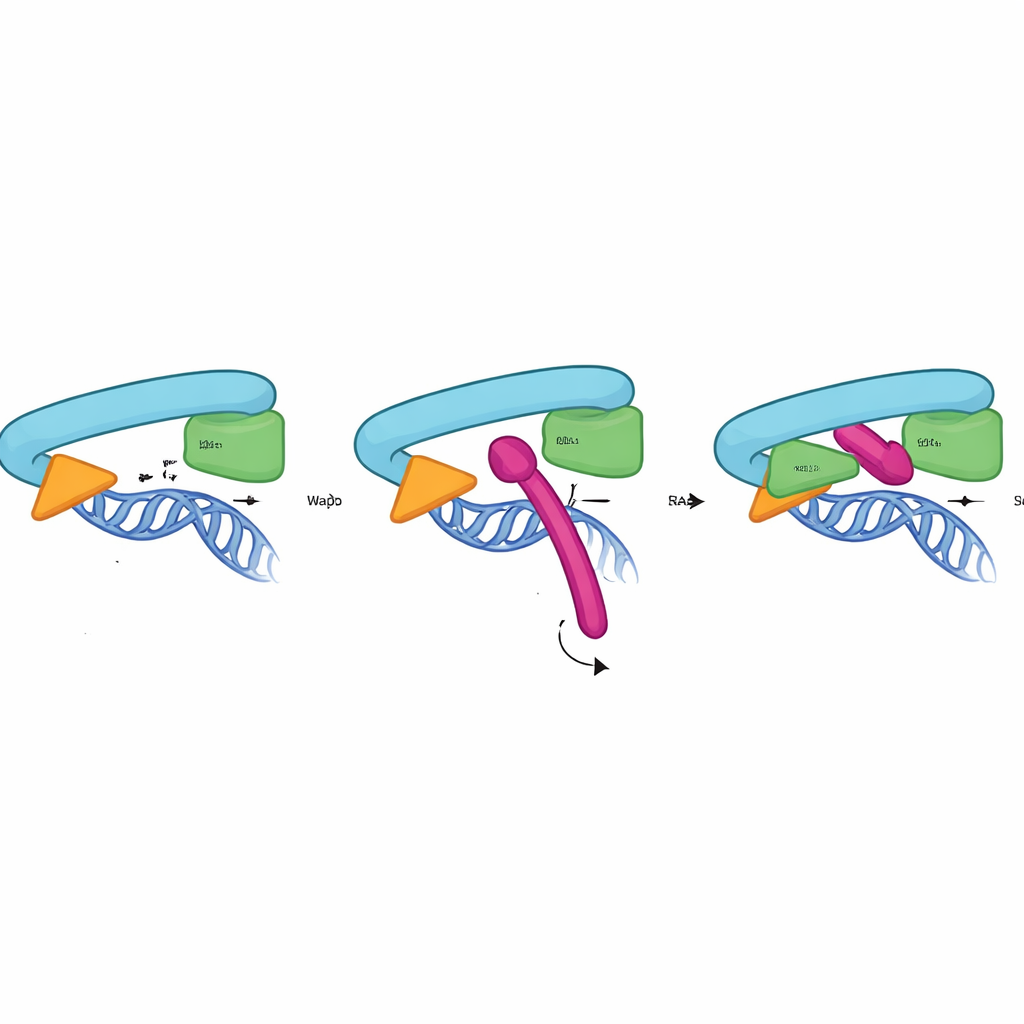
Cómo la cerradura de Sororin sujeta el anillo
Para comprender cómo funciona la CTR a nivel atómico, los investigadores combinaron pruebas bioquímicas, mutagénesis y predicciones estructurales de AlphaFold3. Mostraron que la CTR se une directa y específicamente a la interfaz entre RAD21 y la región de cabeza de SMC3: el mismo sitio que forma la puerta de salida del ADN. Esta interacción no requiere Pds5 ni la subunidad SA2 y ocurre solo en la cohesina unida a la cromatina, alcanzando su punto máximo en la fase en que la cohesión debe mantenerse estable. El modelo estructural sugiere que una hélice corta dentro de la CTR encaja en un bolsillo formado por RAD21 y SMC3, empleando un cúmulo de residuos conservados, hidrofóbicos y cargados negativamente para anclarse. Mutar solo algunos de estos puntos de contacto en Sororin, RAD21 o SMC3 fue suficiente para abolir la unión y provocar la separación prematura de las hermanas, confirmando que esta interfaz actúa como una cerradura física de la puerta.
Un interruptor temporizado para la separación cromosómica
Las células también deben liberar la cohesina en el momento adecuado. El equipo descubrió que Sororin lleva un sitio de control próximo, un único aminoácido (S145) que es fosforilado por la quinasa mitótica Aurora B. Esta modificación rompe específicamente el agarre de Sororin sobre Pds5, permitiendo que Wapl vuelva a participar y favorezca la eliminación de cohesina de los brazos cromosómicos, pero deja en gran medida intacto el contacto de la CTR con la puerta RAD21–SMC3. En los centrómeros, otro factor, Sgo1, recluta una fosfatasa que elimina este fosfato, preservando la unión Sororin–Pds5 y el bloqueo fuerte de la puerta hasta el corte final de RAD21 en anafase. Ensayos genéticos mostraron que cuando se elimina Wapl, los defectos de cohesión causados por la interrupción de la interacción CTR–puerta desaparecen, subrayando que la función esencial de Sororin es proteger la puerta de salida específicamente frente a la apertura impulsada por Wapl.
Qué implica esto para la estabilidad genómica y la enfermedad
En conjunto, estos hallazgos reinterpretan a Sororin como un guardián de doble función: compite con Wapl por el acceso a Pds5 y a la vez cierra directamente la puerta de salida del ADN de la cohesina mediante su pequeña CTR. Este control dual asegura que la cohesión sea fuerte donde se necesita, especialmente en los centrómeros, pero que pueda retirarse de los brazos cromosómicos en la mitosis temprana para permitir una compactación y desenredo correctos. Dado que las mutaciones en la cohesina y sus reguladores son comunes en trastornos del desarrollo y cánceres, la superficie de contacto mapeada con precisión entre Sororin, RAD21 y SMC3 destaca una nueva vulnerabilidad en la que pequeños cambios pueden desestabilizar los cromosomas —y un posible objetivo para futuras terapias que busquen ajustar cuán firmemente se sujeta nuestro material genético.
Cita: Chen, Q., Yuan, X., Shi, M. et al. Sororin locks the DNA-exit gate of cohesin to preserve sister-chromatid cohesion. Nat Commun 17, 2284 (2026). https://doi.org/10.1038/s41467-026-70484-2
Palabras clave: cohesión de las cromátidas hermanas, complejo cohesina, Sororin, segregación cromosómica, estabilidad genómica