Clear Sky Science · es
Un mecanismo adaptador multivalente impulsa la importación nuclear de los proteasomas
Cómo las células mantienen su sala de control limpia
Dentro de cada célula, el núcleo funciona como un centro de mando, repleto de ADN y de las proteínas que regulan qué genes se activan o se silencian. Para mantener esa sala de control en buen estado, las células dependen de potentes “trituradoras” moleculares llamadas proteasomas, que destruyen proteínas desgastadas o defectuosas. Este artículo revela cómo las células gestionan la tarea, sorprendentemente compleja, de transportar estos voluminosos proteasomas a través de las estrechas puertas hacia el núcleo en el momento adecuado.
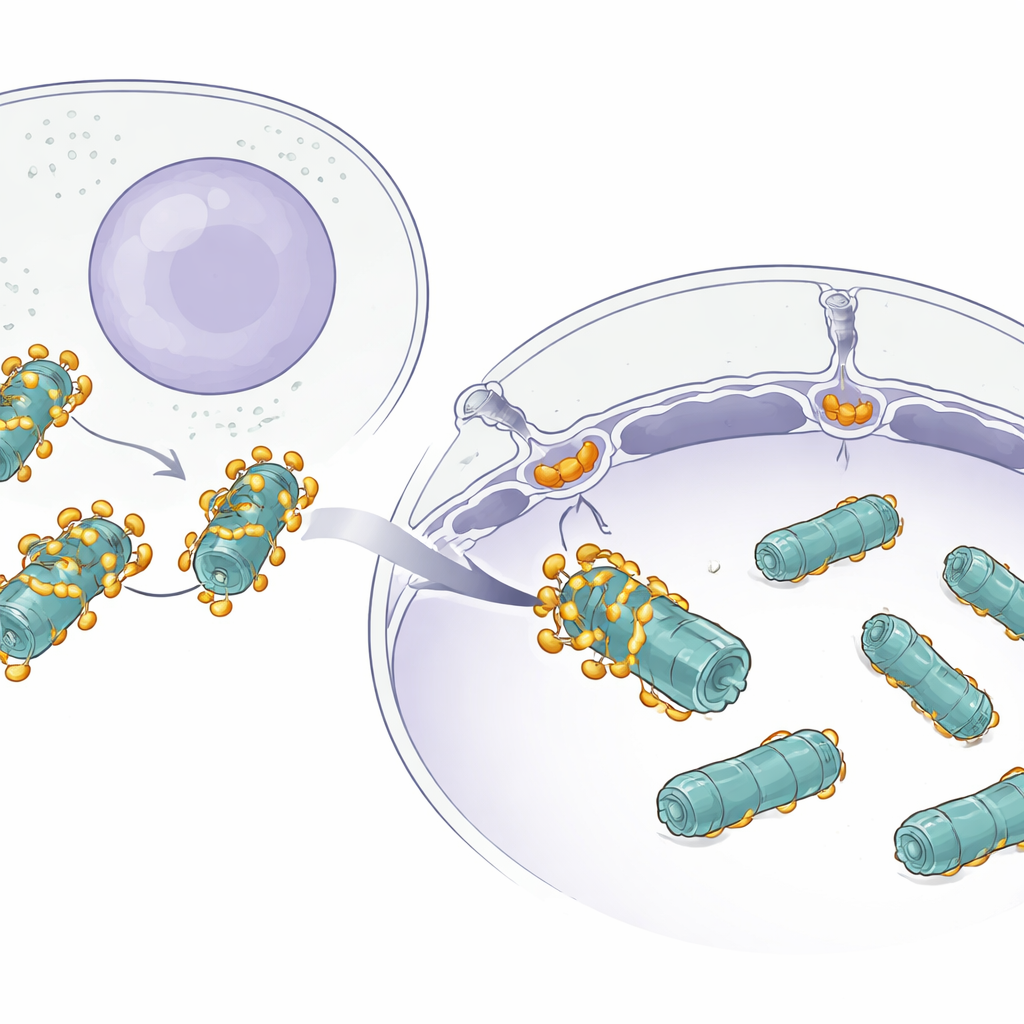
El desafío de mover máquinas grandes
Muchas proteínas se desplazan silenciosamente entre el citoplasma y el núcleo a través de enormes puertas en la envoltura nuclear conocidas como poros nucleares. Las proteínas pequeñas pueden pasar con facilidad, pero las máquinas grandes como los proteasomas son distintas: son complejos masivos con forma de barril formados por muchas piezas. Aunque ya se sabía que las cargas pequeñas usan proteínas auxiliares llamadas importinas para entrar en el núcleo, no estaba claro cómo algo tan grande como un proteasoma logra atravesar eficazmente sin atascarse o perderse.
Un ayudante que cambia de forma cobra protagonismo
El equipo se centró en una proteína pequeña y flexible llamada AKIRIN2, que trabajos previos habían mostrado como absolutamente necesaria para que los proteasomas entren en el núcleo. Las células que se dividen sin AKIRIN2 no pueden introducir proteasomas en el núcleo y mueren rápidamente porque las proteínas nucleares dañadas se acumulan. Usando una combinación de métodos —incluido un extenso escaneo mutacional que alteró casi todos los aminoácidos de AKIRIN2, microscopía crioelectrónica de alta resolución para ver el complejo en 3D y pruebas bioquímicas cuidadosas— los investigadores mapearon qué partes de AKIRIN2 importan y qué hacen. Descubrieron un segmento en forma de “ala” hasta entonces no apreciado y una región en hélice que permite a AKIRIN2 formar dímeros (pares), ambos con papeles clave en sujetar a los proteasomas y a otros socios.
Construir un mango de transporte con muchas manos
Las imágenes estructurales revelaron que múltiples copias de AKIRIN2 se agrupan en un lado del proteasoma como muchas manos agarrando el mismo objeto. Cada par de AKIRIN2 se une a puntos específicos en la superficie externa del proteasoma mientras interactúa con distintas importinas. Una importina, IPO9, se aferra a AKIRIN2 de una manera inusual, envolviéndola y cambiando de forma a medida que se ensambla el complejo. Otras importinas, trabajando también en pares, reconocen señales cortas de entrada nuclear que porta AKIRIN2. Dado que varios AKIRIN2 se sitúan sobre el mismo proteasoma, colectivamente muestran muchas de estas señales a la vez. Este “mango” multivalente permite que una multitud de importinas se enganchen simultáneamente, dando al enorme proteasoma suficiente fuerza de tracción para moverse a través del poro nuclear.
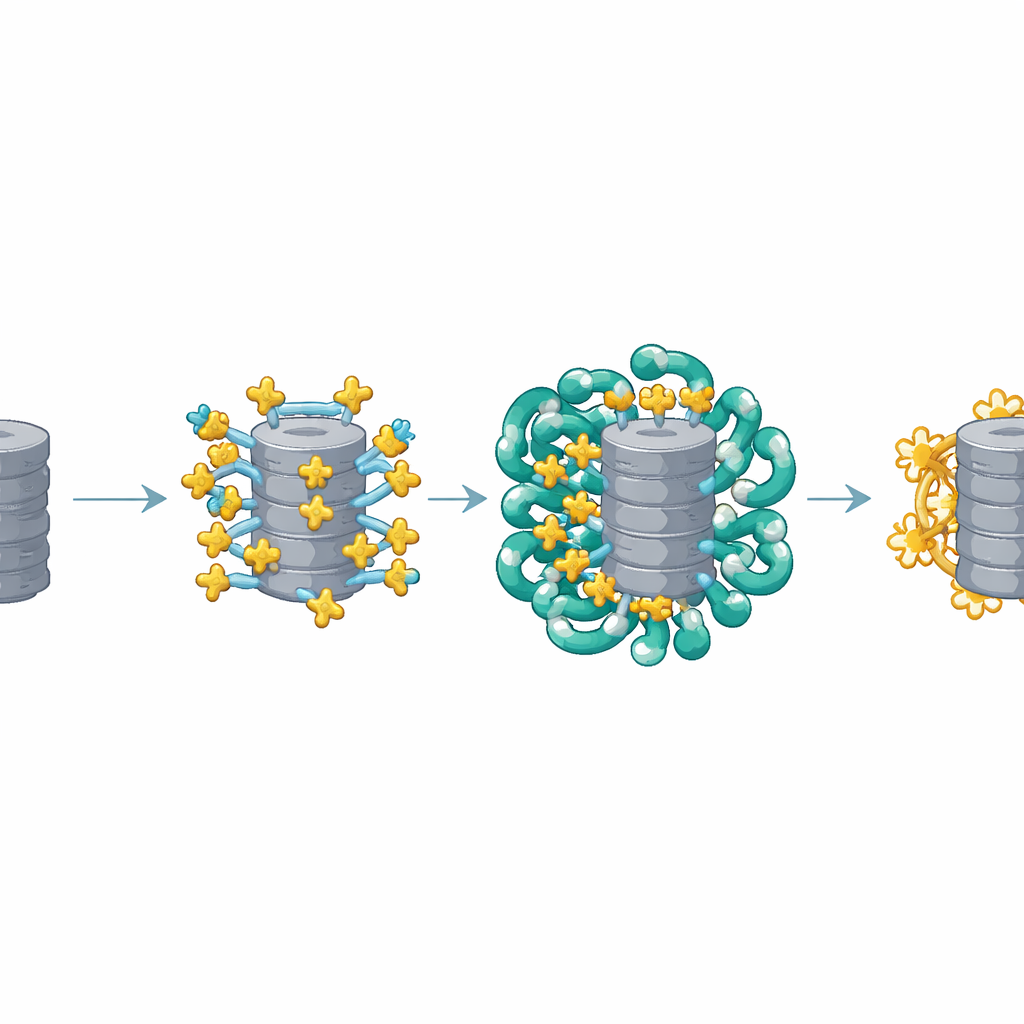
Un ciclo inteligente de ensamblaje y liberación
Experimentos de reconstitución en un sistema simplificado de tubo de ensayo mostraron que los proteasomas solo entran en los núcleos de forma eficiente cuando tanto AKIRIN2 como las importinas están presentes simultáneamente. Cuanto más AKIRIN2, más fuerte y rápida es la importación. Una vez dentro del núcleo, la maquinaria de transporte debe retirarse para que los proteasomas puedan actuar. Aquí la célula utiliza una liberación en dos pasos. Una pequeña proteína conmutadora llamada Ran, en su forma ligada a GTP, desprende la mayor parte de las importinas de su carga. AKIRIN2 y parte de IPO9 se mantienen con más persistencia, pero el núcleo tiene un respaldo: los proteasomas nucleares degradan directamente a AKIRIN2, sin el habitual marcado con ubiquitina. Medidas a lo largo del ciclo celular muestran que AKIRIN2 alcanza su máximo durante la división celular, cuando se forman nuevos núcleos, y luego disminuye en el primer estadio G1, coherente con esta limpieza dirigida.
Por qué esto importa más allá de una proteína
En conjunto, estos hallazgos muestran que AKIRIN2 actúa como un andamio flexible que reúne muchas importinas sobre un único proteasoma, convirtiendo un problema de transporte difícil en un esfuerzo de grupo coordinado. Al agrupar las señales de entrada nuclear en un adaptador en lugar de en el propio proteasoma, las células pueden controlar cuándo y con qué intensidad máquinas grandes se introducen en el núcleo sin rediseñar esas máquinas. Estrategias basadas en adaptadores similares parecen utilizarse para otros grandes ensamblajes de proteínas, e incluso los virus explotan trucos relacionados para invadir el núcleo. Comprender esta lógica transportadora generaliza nuestra visión de cómo las células mantienen el equilibrio proteico en su compartimento más sensible y podría, eventualmente, apuntar a nuevas formas de intervenir cuando la degradación de proteínas o la función nuclear fallan.
Cita: Brunner, H.L., Kalis, R.W., Grundmann, L. et al. A multivalent adaptor mechanism drives the nuclear import of proteasomes. Nat Commun 17, 2359 (2026). https://doi.org/10.1038/s41467-026-69162-0
Palabras clave: transporte nuclear, proteasoma, AKIRIN2, importina, degradación de proteínas