Clear Sky Science · es
Roles distintos de los subtipos de la capa cortical 5 en el aprendizaje asociativo
Cómo el cerebro aprende que un toque anuncia una recompensa
Imagínese aprender que un ligero golpecito en la muñeca significa que viene el postre. Su cerebro debe vincular un simple contacto con una recompensa futura. Este estudio examina el cerebro del ratón para ver cómo dos tipos distintos de células nerviosas trabajan en conjunto para formar esas asociaciones, revelando cómo el aprendizaje cotidiano se divide entre “lo que ocurrió” y “lo que eso significa para mí”. 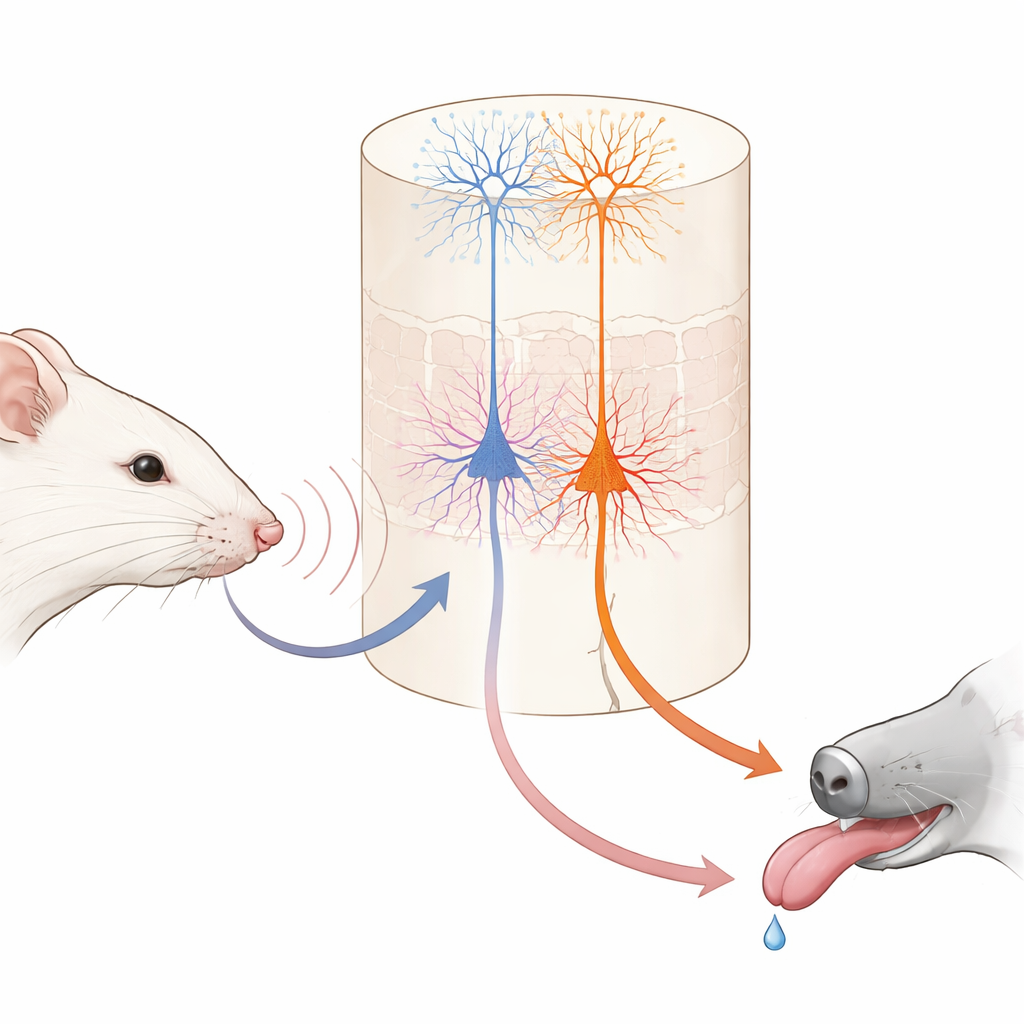
Un juego de vibrisas sencillo para estudiar el aprendizaje
Los investigadores entrenaron ratones con la cabeza fijada en un juego sencillo usando sus vibrisas faciales. En cada ensayo, una sola vibrisa era vibrada a una de dos velocidades. Una vibración iba seguida, tras un breve retraso, de una pequeña gota de agua; la otra nunca se recompensaba. Al principio, los ratones lamían anticipadamente en ambos tipos de ensayo, sin saber todavía qué sensación en la vibrisa señalaba el agua. Tras varios días, aprendieron gradualmente a lamer temprano solo cuando ocurría la vibración “buena” y a contenerse cuando aparecía la no recompensada. Cuando los científicos inactivaron temporalmente el área sensorial primaria durante el entrenamiento, este aprendizaje desapareció en gran medida, lo que demuestra que esta región sensorial es importante para construir la asociación, aunque los ratones expertos podían después realizar la tarea sin ella.
Dos tipos de células de salida con trabajos muy distintos
Dentro de esta área táctil, una capa profunda conocida como la capa 5 contiene dos tipos principales de células nerviosas de salida. Un grupo, llamado aquí células IT, envía señales a otras regiones corticales en ambos hemisferios. El otro grupo, células ET, envía señales principalmente hacia estructuras subcorticales implicadas en la acción y la recompensa. Usando ratones genéticamente modificados e imagen de dos fotones de alta resolución, los autores pudieron monitorizar de forma selectiva la actividad de cada tipo celular a través de las puntas de sus largas y ramificadas dendritas. Antes del aprendizaje, las células IT ya respondían con fuerza y fiabilidad a las vibraciones de las vibrisas, y su actividad conjunta podía distinguir con precisión las dos velocidades de vibración. Las células ET, en contraste, respondían de forma más débil y menos consistente a los estímulos, ofreciendo solo una lectura imprecisa de qué vibración había ocurrido.
Sensaciones estables frente a expectativas crecientes
A medida que los ratones aprendían, las células IT se comportaron como informantes fiables. Sus respuestas se mantuvieron estrechamente ligadas al momento del movimiento de la vibrisa y cambiaron poco día a día. Continuaron codificando qué vibración había ocurrido, independientemente de si predecía una recompensa. Las células ET, sin embargo, transformaron su comportamiento. En lugar de dispararse simplemente al inicio del estímulo, su actividad aumentó gradualmente durante la vibración y el breve retraso, alcanzando un pico alrededor del momento esperado de entrega del agua. Esta subida creció en paralelo con el lamido anticipatorio de los animales y llegó a predecir mejor si un ensayo acabaría en lamido que a reportar el estímulo exacto. Las células ET individuales entraron y salieron del grupo activo a lo largo de los días, pero a nivel poblacional el patrón se volvió más consistente, lo que sugiere un código flexible pero convergente para la expectativa de recompensa. 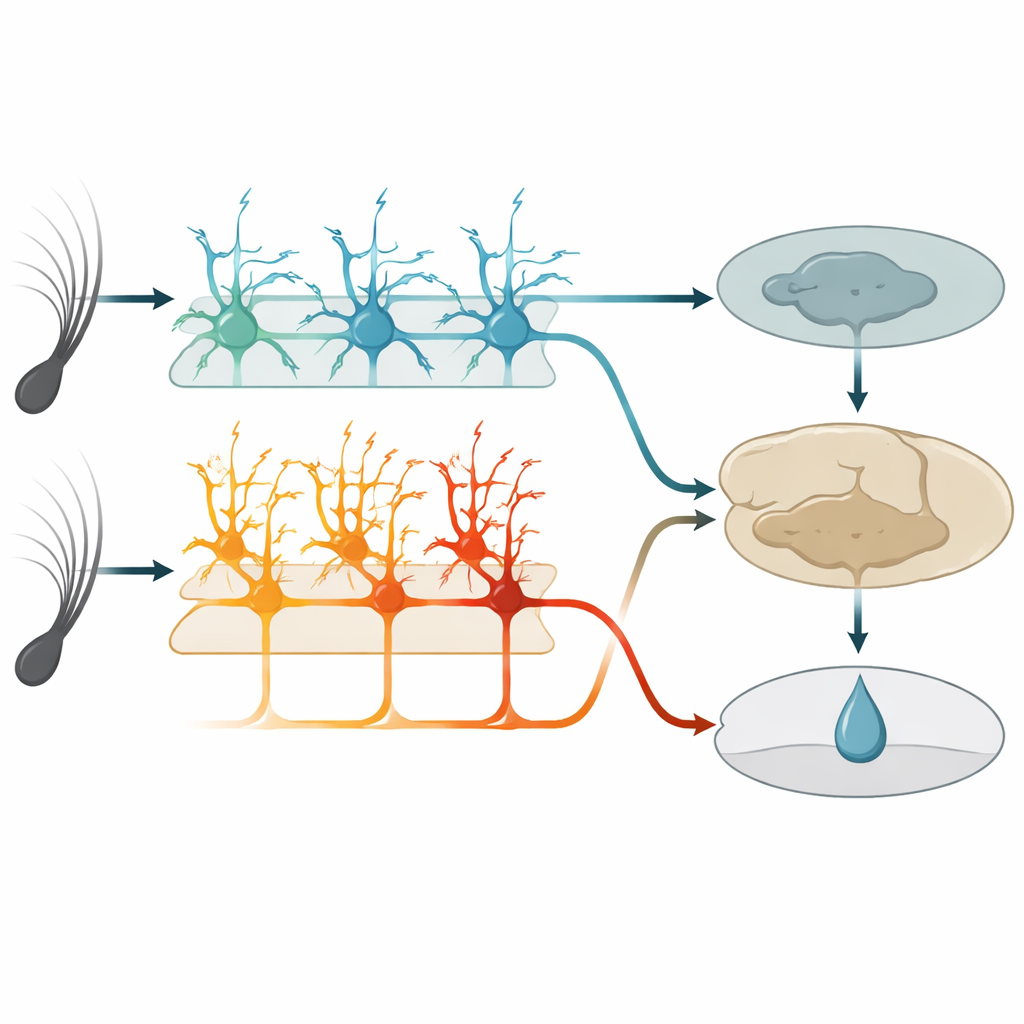
Apagar cada tipo celular revela una división del trabajo
Para testar la función, el equipo usó herramientas quimio-genéticas para amortiguar selectivamente ya fueran las células IT o las ET durante el entrenamiento. Cuando se silenciaron las células IT, los ratones mostraron menos lamidos anticipatorios en general y no consiguieron construir una diferencia clara entre las vibraciones recompensadas y las no recompensadas. Cuando se silenciaron las células ET, los ratones hicieron lo contrario: lamían en exceso en ambos tipos de vibración, especialmente en la no recompensada, y les costó refinar su comportamiento aunque seguían lamiendo vigorosamente. Silenciar cualquiera de los grupos después de que la tarea fue dominada ya no perjudicó el rendimiento, lo que implica que una vez que otras regiones cerebrales han almacenado la asociación, esta área sensorial y sus salidas de la capa 5 dejan de ser críticas para ejecutar la respuesta aprendida.
Un modelo de aprendizaje que refleja el comportamiento cerebral
Los autores construyeron un modelo computacional al estilo de aprendizaje por refuerzo para interpretar estos hallazgos. En el modelo, una red tipo IT proporciona representaciones sensoriales estables que ayudan a estimar el “valor” de cada estímulo—qué probabilidad hay de que vaya seguida por recompensa. Una vía tipo ET transmite este valor predicho a un circuito aguas abajo que lo compara con la recompensa real, generando un error de predicción que ajusta las estimaciones de valor futuras. Bloquear las vías tipo IT o ET en el modelo reprodujo los fracasos de aprendizaje distintos observados en los ratones: sin entrada tipo IT, el aprendizaje era lento y débil para ambos estímulos; sin salida tipo ET, se producía un aprendizaje inicial pero el sistema no lograba reducir adecuadamente las respuestas al indicio no recompensado. El modelo también capturó cómo, con el tiempo, regiones no sensoriales podían asumir la ejecución, coherente con los experimentos.
Qué significa esto para el aprendizaje cotidiano
En términos sencillos, este estudio sugiere que cuando aprendemos que una vista, un sonido o un toque predice algo bueno o malo, distintos conjuntos de neuronas corticales profundas comparten el trabajo. Un conjunto mantiene un registro fiel de “lo que ocurrió” en el mundo, mientras que otro gradualmente llega a señalar “qué espero que ocurra a continuación” y ayuda a afinar el comportamiento comparando esas expectativas con la realidad. Juntos forman un puente entre la sensación cruda y acciones flexibles basadas en la experiencia, ofreciendo una imagen más clara de cómo el cerebro sostiene los hábitos, el aprendizaje por recompensas y, posiblemente, cómo estos procesos se ven alterados en la enfermedad.
Cita: Moberg, S., Garibbo, M., Mazo, C. et al. Distinct roles of cortical layer 5 subtypes in associative learning. Nat Commun 17, 2648 (2026). https://doi.org/10.1038/s41467-026-68307-5
Palabras clave: aprendizaje asociativo, corteza somatosensorial, neuronas de la capa 5, expectativa de recompensa, aprendizaje por refuerzo