Clear Sky Science · en
MYC modulates TOP2A diffusion to promote substrate detection and activity
Why this matters for our DNA
Every time a cell reads its genes, it twists and tangles its DNA. If these knots are not quickly removed, they can stall gene activity and damage the genome. This study uncovers how a well‑known cancer‑linked protein called MYC speeds up the movement of a DNA “untangling” enzyme, TOP2A, inside the cell nucleus. By learning how MYC supercharges this enzyme, researchers point to new ways of selectively slowing down runaway gene activity in cancer cells while sparing normal cells.
DNA knots and the cell’s untangling tools
DNA inside our cells is packed into a tiny space, yet it must constantly be read, copied, and repaired. These activities twist the double helix and create supercoils and crossings, much like kinks in an over‑wound phone cord. Specialized enzymes called topoisomerases solve this problem by cutting DNA strands, passing them around one another, and sealing them back up. TOP2A, one member of this family, makes temporary cuts in both strands of the DNA helix to remove serious tangles. At the same time, MYC acts as a powerful booster of gene activity in many cancers, driving transcription so hard that it generates extra torsional stress on DNA. The big question addressed here is how MYC prevents its own burst of gene activity from choking on the DNA knots it creates.
A moving balance inside the nucleus
The researchers first mapped where TOP2A sits in the nucleus of human cancer cells. They found that TOP2A constantly shuttles between two main zones: the nucleolus, a dense region where ribosomes are made, and scattered transcription hubs where many genes are actively read. This motion forms a dynamic equilibrium, with TOP2A able to relocate rapidly when DNA stress increases. When the scientists artificially increased supercoiling, for example by disabling a related enzyme or blocking TOP2A’s ability to reset after cutting, TOP2A quickly left the nucleolus and accumulated throughout the rest of the genome, where it could act on stressed DNA. Importantly, this shift correlated with more TOP2A molecules being chemically “caught in the act” on DNA, showing that relocation reflects true working engagement, not just passive drifting.
Three modes of motion for TOP2A
Using single‑molecule tracking in living cells, the team showed that TOP2A does not all move the same way. Instead, it exists in three diffusive states. A “bound” state hardly moves and reflects TOP2A clamped onto chromatin. A “slow” state wanders within tiny regions that match the size and location of transcription condensates—droplet‑like hubs where transcription machinery concentrates. A “fast” state zips more freely through the nucleoplasm. The nucleolus contains mostly slow and bound TOP2A, while the rest of the nucleus has a mix of all three. When TOP2A was trapped on DNA by a drug, the bound fraction grew at the expense of the faster states. Together, these observations suggest that TOP2A roams quickly to search for trouble spots, briefly samples transcription condensates, and then becomes fully engaged on DNA when it finds a tangle.
How MYC makes TOP2A move faster
The key discovery is that MYC acts as an “accelerant” for TOP2A. When MYC was rapidly removed from cells, TOP2A’s diffusion slowed in both the fast and slow pools, but its motion in the nucleolus—where MYC is scarce—did not change. Biochemical fractionation confirmed that, without MYC, TOP2A tends to form larger molecular assemblies. In test‑tube experiments, purified TOP2A can form dense droplets that resemble protein condensates. Adding MYC made these droplets smaller and less prone to sink, consistent with weaker self‑clustering and more mobile complexes. This effect did not require another topoisomerase partner, TOP1, although TOP1 can join the same complexes. In short, MYC limits how many TOP2A molecules clump together, shrinking the average complex size and allowing each TOP2A molecule to diffuse more rapidly through the nuclear environment.
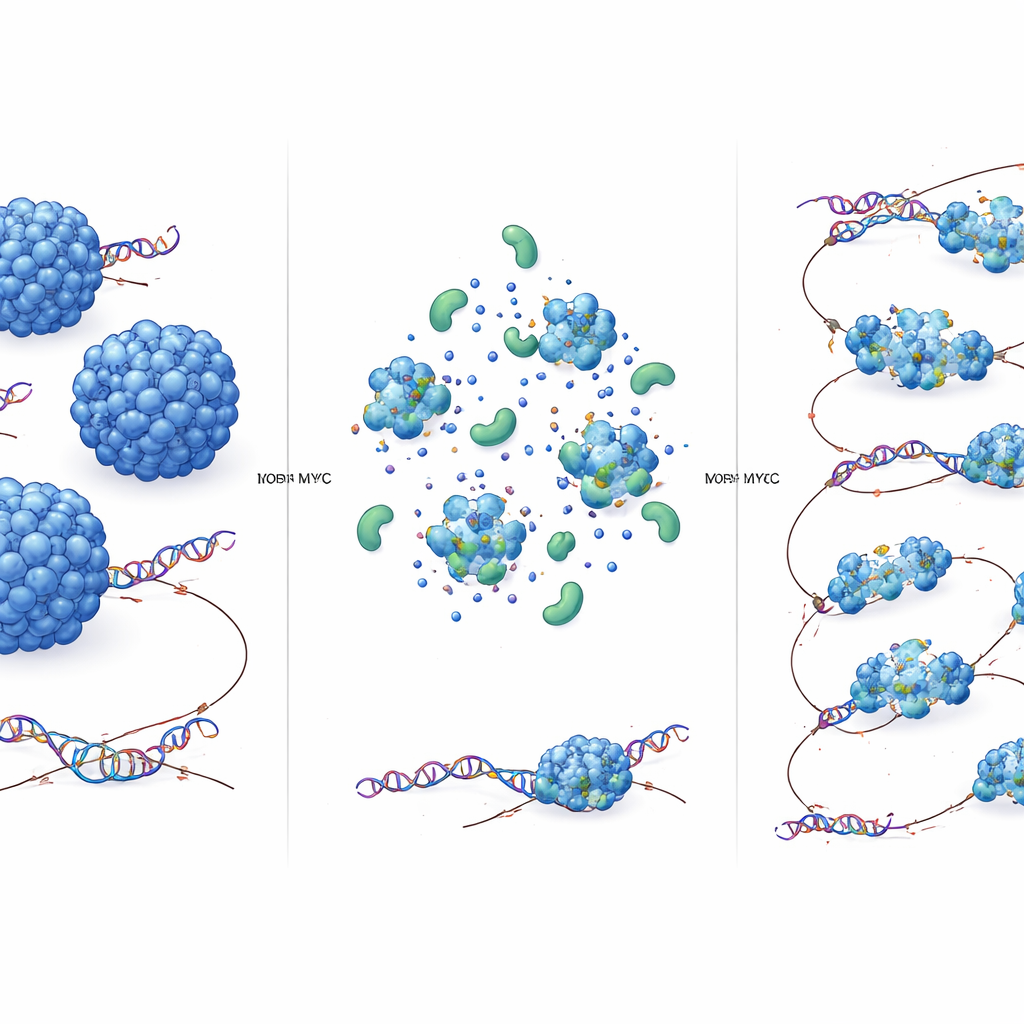
Faster search, more DNA untangling
Does this speed boost matter for function? Using a specialized optical tweezers setup, the authors built single DNA crossovers—tiny mimics of DNA knots—and watched fluorescent TOP2A bind to them. When MYC was present, TOP2A visited these crossovers more often, indicating improved substrate detection. In cells, a genome‑wide assay that captures TOP2A molecules covalently attached to DNA showed strong TOP2A activity at the beginnings and ends of highly expressed genes. Rapid MYC depletion substantially reduced these active TOP2A–DNA complexes, even though overall transcription levels barely changed in that time window. This means MYC directly enhances TOP2A’s ability to find and act on tangled DNA rather than simply raising gene expression.
What this means for cancer and future treatments
Putting these pieces together, the study proposes a simple but powerful idea: by keeping TOP2A complexes relatively small, MYC makes them diffuse faster, find DNA tangles more efficiently, and work harder at sites of intense transcription. In healthy cells, this coupling helps maintain smooth gene activity. In MYC‑driven cancers, however, the same mechanism may turbocharge DNA untangling in ways that let tumor cells tolerate extreme transcriptional stress. Targeting the MYC–TOP2A partnership, or the way MYC reshapes TOP2A condensates, could therefore offer a route to selectively weaken cancer cells’ ability to manage DNA strain without shutting down essential processes in normal tissues.
Citation: Cameron, D.P., Jackson, K., Loffreda, A. et al. MYC modulates TOP2A diffusion to promote substrate detection and activity. Nat Commun 17, 2527 (2026). https://doi.org/10.1038/s41467-026-69232-3
Keywords: MYC, TOP2A, DNA topology, transcription condensates, cancer biology