Clear Sky Science · de
Der Perforansweg und CA3‑Schaffer‑Collateral‑Afferenzen koordinieren sich, um räumliches Lernen zu regulieren
Warum es wichtig ist, den Weg zu finden
Ob eine Maus eine versteckte Plattform sucht oder ein Mensch sich in einer neuen Stadt zurechtfindet: Das Gehirn wandelt fortlaufend Seh‑, Hör‑ und Eigenbewegungsinformationen in eine interne Karte um. Diese Studie untersucht, wie zwei zentrale Hirnregionen, der entorhinale Kortex und der Hippocampus, zusammenarbeiten, um diese Fähigkeit zu unterstützen. Indem die Forschenden die Aktivität winziger Nervenfasern live beobachteten und spezifische Bahnen bei Mäusen gezielt stimulierten oder stilllegten, zeigen sie, wie diese Bereiche koordinieren, um räumliche Erinnerungen zu bilden und zu stabilisieren – Erkenntnisse, die möglicherweise helfen können, Gedächtnisstörungen besser zu verstehen und zu behandeln.
Zwei Schaltzentralen des inneren GPS
Hippocampus und der benachbarte entorhinale Kortex liegen tief im Temporallappen und bilden den Kern unseres Navigationssystems. Innerhalb des Hippocampus verbindet ein innerer Schaltkreis einen Bereich namens CA3 mit einem anderen namens CA1 über Fasern, die als Schaffer‑Collateral bezeichnet werden. Gleichzeitig sendet der entorhinale Kortex einen separaten Informationsstrom direkt in CA1 über den so genannten Perforansweg. Die Autorinnen und Autoren wollten herausfinden, wie diese beiden Eingangsströme zusammenwirken, wenn ein Tier die Struktur eines Raumes erlernt, und wie Veränderungen an ihren Verbindungen – allgemein als „Plastizität“ bezeichnet – den Aufbau einer stabilen räumlichen Karte unterstützen.
Das Lernen Faser für Faser beobachten
Um diesen Prozess in verhaltenden Tieren zu verfolgen, verwendete das Team Fiber‑Photometry, eine Technik, die Nervenaktivität über Leuchtsignale fluoreszierender Proteine anzeigt. Sie konstruierten Mäuse so, dass CA3‑Neurone, die zu CA1 projizieren, bei Aktivität aufleuchteten, und trainierten die Tiere im Morris‑Wassertest, einem klassischen Paradigma, in dem eine Maus die versteckte Lage einer kleinen Plattform in einem Becken lernen muss. In frühen Versuchen waren die CA3–CA1‑Fasern während der Suche stark aktiv; mit zunehmender Gewandtheit und schnelleren Auffindzeiten nahm diese Aktivität schrittweise ab. Dieses Muster deutet darauf hin, dass diese Verbindungen besonders während der Erstkodierung der Raumstruktur engagiert sind und sich nach Etablierung der Erinnerung in eine effizientere, stabilere Repräsentation einpendeln.
Wie upstream‑Input Lernen verstärkt oder abschwächt
Die Forschenden fragten als Nächstes, wie Signale aus dem entorhinalen Kortex diesen hippocampalen Schaltkreis beeinflussen. Mit lichtempfindlichen Proteinen aktivierten sie entorhinale Neurone, während sie von den CA3–CA1‑Fasern aufzeichneten, und zeigten, dass die Anregung von Eingängen aus dem medialen entorhinalen Kortex die Aktivität dieser hippocampalen Verbindungen zuverlässig erhöhte. Umgekehrt schwächte eine chemogenetische Dämpfung der entorhinalen Signale, die während des Wassertest‑Trainings in CA1 eintrafen, die CA3–CA1‑Aktivität und die Mäuse lernten die Plattformlage langsamer und weniger präzise. In einem ergänzenden Experiment zeichneten die Autorinnen und Autoren direkt von entorhinalen Neuronen auf, die zu CA1 projizieren, und beobachteten, dass deren Aktivität über die Trainingstage anstieg – parallel zur Leistungsverbesserung der Tiere. Zusammen deuten diese Befunde darauf hin, dass starke, zeitlich gut abgestimmte entorhinale Signale nötig sind, um hippocampale Schaltkreise für die Navigation richtig einzustellen.
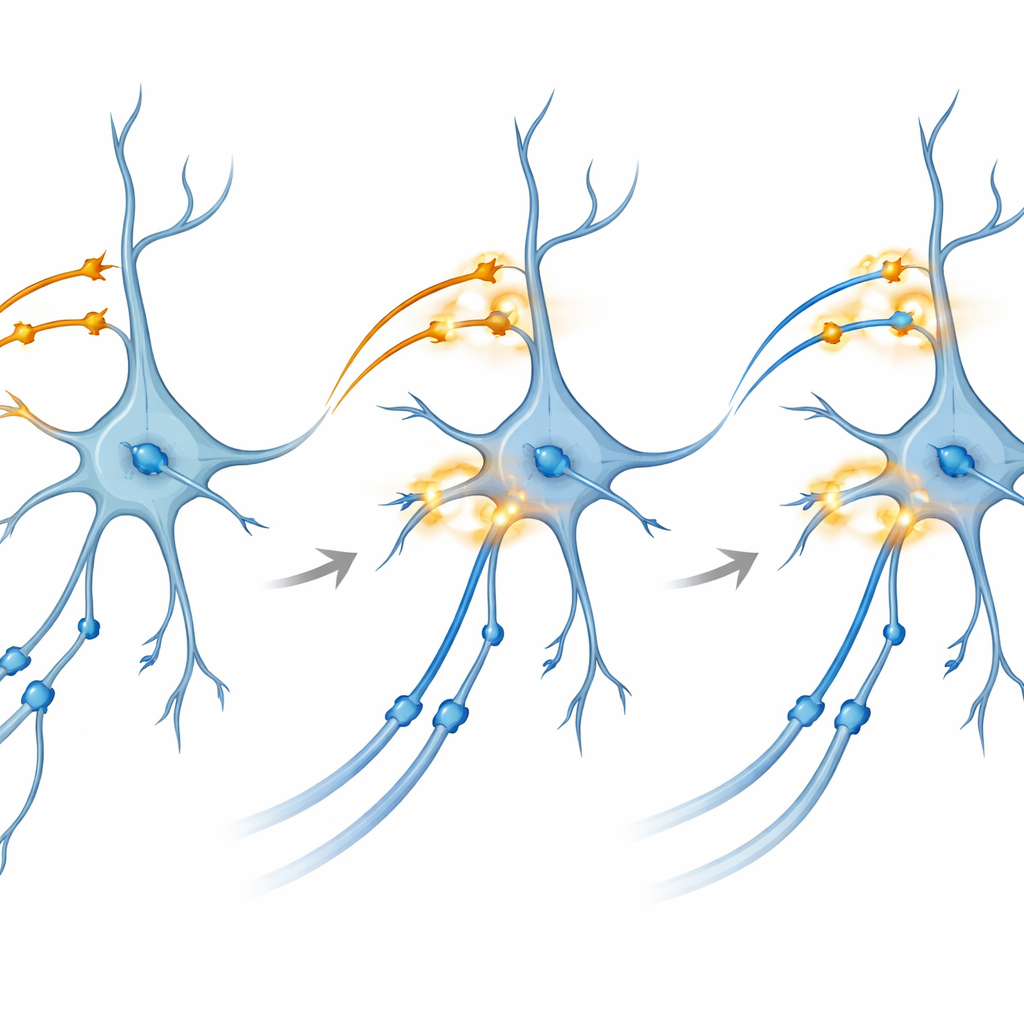
Warum Verbindungen stärker werden: ein genauerer Blick
Um den zugrunde liegenden Mechanismus zu untersuchen, arbeitete das Team mit Gehirnschnitten, die in einer Nährlösung lebendig gehalten wurden. Dort konnten sie lichtaktivierbare Proteine in CA3 und in entorhinalen Eingängen zu CA1 getrennt kontrollieren. Überraschenderweise erzeugte die alleinige Anwendung von Theta‑Burst‑Lichtstimulation – Muster, die natürliches rhythmisches Feuern nachahmen – auf einen der beiden Wege nicht zuverlässig eine langanhaltende Potenzierung (LTP), die anhaltende Verstärkung von Synapsen, die als Grundlage des Gedächtnisses gilt. Wenn sie jedoch einen sorgfältig zeitlich abgestimmten, dualfarbigen Burst einsetzten, der beide Pfade gleichzeitig in CA1 koaktivierte, zeigten die CA3–CA1‑Verbindungen eine robuste und lang anhaltende Verstärkung. Die Blockade von NMDA‑Rezeptoren oder bestimmten Calciumkanälen, zwei bekannten molekularen Schaltern für Plastizität, verhinderte diesen Effekt und verband das Phänomen mit klassischen biochemischen Pfaden der Gedächtnisbildung.
Folgerungen für Gedächtnis und Krankheit
Insgesamt zeichnet die Studie das Bild räumlichen Lernens als eine koordinierte Choreografie zwischen eintreffenden entorhinalen Signalen und der internen hippocampalen Verschaltung. Entorhinale Eingänge liefern nicht nur Informationen über Position und Kontext; sie fungieren auch als kraftvoller Regelknopf, der die Aktivität in CA3–CA1‑Fasern verstärken oder abschwächen kann und bei Koaktivierung die langfristige Verstärkung dieser Synapsen antreibt. Für eine interessierte Leserschaft ist die Kernaussage, dass das Gehirn‑"GPS" nicht auf eine einzelne Region angewiesen ist, sondern auf eine Partnerschaft, in der eine Region die Szene setzt und eine andere die Details festschreibt. Störungen dieser Partnerschaft – etwa durch Alterung, Verletzung oder neurodegenerative Erkrankungen – könnten erklären, warum Menschen sich in vertrauten Umgebungen verirren, und die gezielte Ansprache dieser Bahnen könnte neue Wege eröffnen, räumliches Gedächtnis zu erhalten oder wiederherzustellen.
Zitation: Huang, F., Temitayo Bello, S., Lau, S.H. et al. The perforant pathway and CA3-Schaffer collateral afferents coordinate to regulate spatial learning. Commun Biol 9, 364 (2026). https://doi.org/10.1038/s42003-026-09577-z
Schlüsselwörter: räumliches Gedächtnis, Hippocampus, entorhinale Kortex, synaptische Plastizität, Navigierung