Clear Sky Science · de
Ein intrakortikales Gehirn‑Maschine‑Interface basierend auf Aktivität des ventralen prämotorischen Kortex beim Makaken
Dem Gehirn beibringen, einen Cursor zu bewegen
Stellen Sie sich vor, Sie lenken einen Computercursor oder einen Roboterarm allein mit Ihren Gedanken, selbst wenn Ihre Muskeln sich nicht mehr bewegen können. Gehirn‑Maschine‑Interfaces (BMIs) wollen genau das ermöglichen, indem sie Gehirnaktivität in Befehle für externe Geräte übersetzen. Die meisten bisherigen Systeme greifen auf eine Hauptbewegungsregion des Gehirns zurück — was aber, wenn diese Region beschädigt ist, etwa bei Schlaganfall oder ALS? Diese Studie untersucht, ob eine nahegelegene Region, die normalerweise mit der Planung von Handbewegungen und dem Beobachten fremder Aktionen in Verbindung gebracht wird, ebenfalls zuverlässig ein BMI steuern kann.
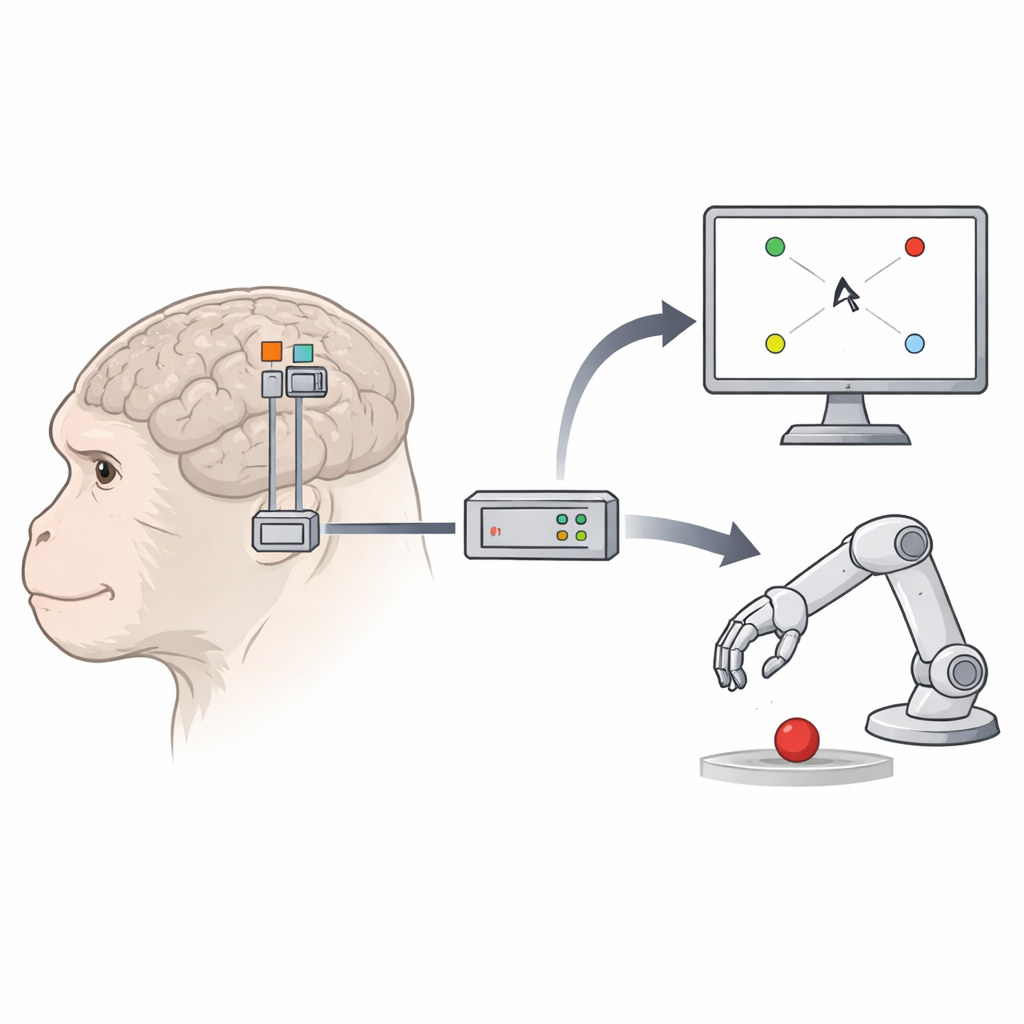
Eine neue Hirnregion kommt ins Spiel
Klassische BMIs lesen hauptsächlich Signale aus dem primären Motorkortex, dem Hirnstreifen, der willkürliche Bewegungen direkt steuert, und aus einer benachbarten Planungsregion, dem dorsalen prämotorischen Kortex. Die Forschenden richteten ihre Aufmerksamkeit auf einen anderen Nachbarn: den ventralen prämotorischen Kortex, speziell eine Zone namens F5c. Bei Affen ist F5c reich an Zellen, die feuern, wenn das Tier greift und erreicht, und sogar, wenn es lediglich Aktionen auf einem Bildschirm beobachtet. Diese Mischung aus Bewegungs‑ und Beobachtungsantworten ließ vermuten, dass F5c gut geeignet sein könnte, einen Cursor oder einen robotischen „Avatar“ zu steuern, ohne dass der Körper sich bewegen muss.
Affen, Mikroelektroden und bewegte Ziele
Zwei Makaken erhielten implantierte 96‑Elektroden‑Gitter an drei Stellen: primärer Motorkortex, dorsaler prämotorischer Kortex und F5c. In täglichen Sitzungen führten die Tiere mehrere visuell einfache, aber verhaltensmäßig anspruchsvolle Aufgaben aus. In einer Aufgabe berührten sie die Bildschirmmitte und reichten dann zu einem von acht äußeren Zielen, während ein kleines Bildschirmquadrat vom Zentrum zum gleichen Ziel wanderte. In einer zweiten Aufgabe hielten sie die Hand still und beobachteten lediglich den Cursor auf dem Weg zu den Zielen. In einer dritten, realitätsnäheren Anordnung wurde der Cursor durch einen 3D‑Roboterarm‑Avatar ersetzt, der in einer virtuellen Szene nach Zielen griff. Über diese Aufgaben hinweg konnte das Team vergleichen, wie gut jede Hirnregion Cursor‑ oder Avatarbewegungen antrieb.
Wie Gehirnsignale zu flüssiger Bewegung wurden
Während einer Trainingsphase folgten Cursor oder Avatar computergenerierten, sanft gekrümmten Pfaden, während die Affen entweder selbst bewegten oder zusahen. Gleichzeitig zeichneten die Elektroden schnelle Ausbrüche neuronaler Aktivität auf. Die Forschenden trainierten dann einen Dekoder — ein mathematisches Werkzeug, das lernt, Muster neuronaler Feuerraten auf die Geschwindigkeiten des Cursors oder Avatars auf dem Bildschirm abzubilden. Um nur die informativsten Kanäle zu erfassen, wählten sie Elektroden aus, deren Aktivität Richtung und Geschwindigkeit der Bewegung nachverfolgte. Sie verwendeten eine Methode, die Gehirnmuster isoliert, die am engsten mit dem Verhalten verknüpft sind, und erweiterten sie durch einen nichtlinearen Schritt, sodass das System komplexere Beziehungen zwischen neuronaler Aktivität und Bewegung erfassen konnte. In der Dekodierungsphase steuerte nicht länger der Computer den Cursor oder Avatar; stattdessen nutzte der Dekoder Live‑Gehirnsignale, aktualisiert alle 50 Millisekunden, um die Onscreen‑Bewegung zu lenken. Der Dekoder wurde periodisch im Hintergrund nachtrainiert, damit er sich an Verschiebungen in den neuronalen Antworten über die Zeit anpassen konnte.

Wie gut schlug sich die „neue“ Region?
Die zentrale Frage war, ob F5c die Leistung der traditionellen Kontrollareale erreichen oder halbwegs annähern kann. Bei beiden Affen hinkte die auf F5c basierende Dekodierung anfangs beim Bewegen des Cursors hinterher, besonders wenn die Tiere nur zusahen und ihre eigene Hand nicht bewegten. Mit fortschreitenden Sitzungen — und sobald mehr Elektroden zuverlässige bewegungsbezogene Signale lieferten — holte F5c jedoch auf. In mehreren Bedingungen erreichte seine Leistung die des primären Motorkortex und übertraf ihn in späteren Sitzungen sogar bei passiver Cursorsteuerung. Bei der Steuerung des Roboter‑Avatars war die Gesamtperformance über alle Regionen hinweg niedriger, doch F5c erlaubte trotzdem eine sinnvolle Kontrolle, insbesondere in Kombination mit einem sanften assistiven Algorithmus, der den Avatar dezent zum Ziel leitete. Wichtig ist, dass Neurone in allen drei Regionen ähnliche Muster richtungsabhängiger Abstimmung und Populationsaktivität sowohl während der computergesteuerten Trainingsphase als auch in der aktiven Steuerungsphase zeigten, wobei nur ein Subset seine bevorzugten Richtungen änderte, als die Affen lernten, das BMI zu bedienen.
Was das für zukünftige Neurotechnologien bedeutet
Für Nicht‑Spezialisten ist die entscheidende Erkenntnis, dass der ventrale prämotorische Bereich F5c — lange vor allem als Planer und Beobachter von Aktionen betrachtet — auch als praktikabler Kontrollknoten für Gehirn‑Maschine‑Interfaces dienen kann. Wenn genügend bewegungsbezogene Signale verfügbar sind, können auf F5c‑Aktivität trainierte Dekoder einen Bildschirmcursor oder einen assistiven Roboter nahezu ebenso gut steuern wie solche, die auf der klassischen Bewegungsregion basieren. Das legt nahe, dass künftige klinische BMIs nicht zwangsläufig auf eine einzige kortikale Region angewiesen sein müssen. Für Menschen mit geschädigtem primärem Motorkortex könnten benachbarte Planungsregionen wie der ventrale prämotorische Kortex einen alternativen Weg bieten, um wieder Kontrolle über digitale Werkzeuge, Prothesen oder Mobilitätshilfen zu erlangen.
Zitation: De Schrijver, S., Garcia Ramirez, J., Iregui, S. et al. An intracortical brain-machine interface based on macaque ventral premotor activity. Sci Rep 16, 8407 (2026). https://doi.org/10.1038/s41598-026-38536-1
Schlüsselwörter: Gehirn‑Maschine‑Interface, motorischer Kortex, prämotorischer Kortex, neuronale Dekodierung, prothetische Steuerung