Clear Sky Science · de
Propriozeptive Grenzdetektoren tragen zur sensomotorischen Steuerung des Drosophila‑Beins bei
Wie Fliegen ihre Schritte sichern
Jedes Mal, wenn ein Bein nach vorne schwingt, besteht die Gefahr des Überstreckens und Stolperns. Von Hauskatzen bis zu Fruchtfliegen verlassen sich Tiere auf verborgene Sinnesorgane, die dem Nervensystem mitteilen, wenn ein Gelenk seinem Anschlag nahe ist. Diese Studie zeigt, wie winzige Sensoren an den Vorderbeinen der Fruchtfliege als eingebaute „Stopwächter“ funktionieren und dem Tier helfen, sein Gewicht zu verlagern, Kollisionen zu vermeiden und eine stabile Haltung zu bewahren – selbst beim schnellen Gehen oder Putzen.
Verborgene Sensoren am Rand der Bewegung
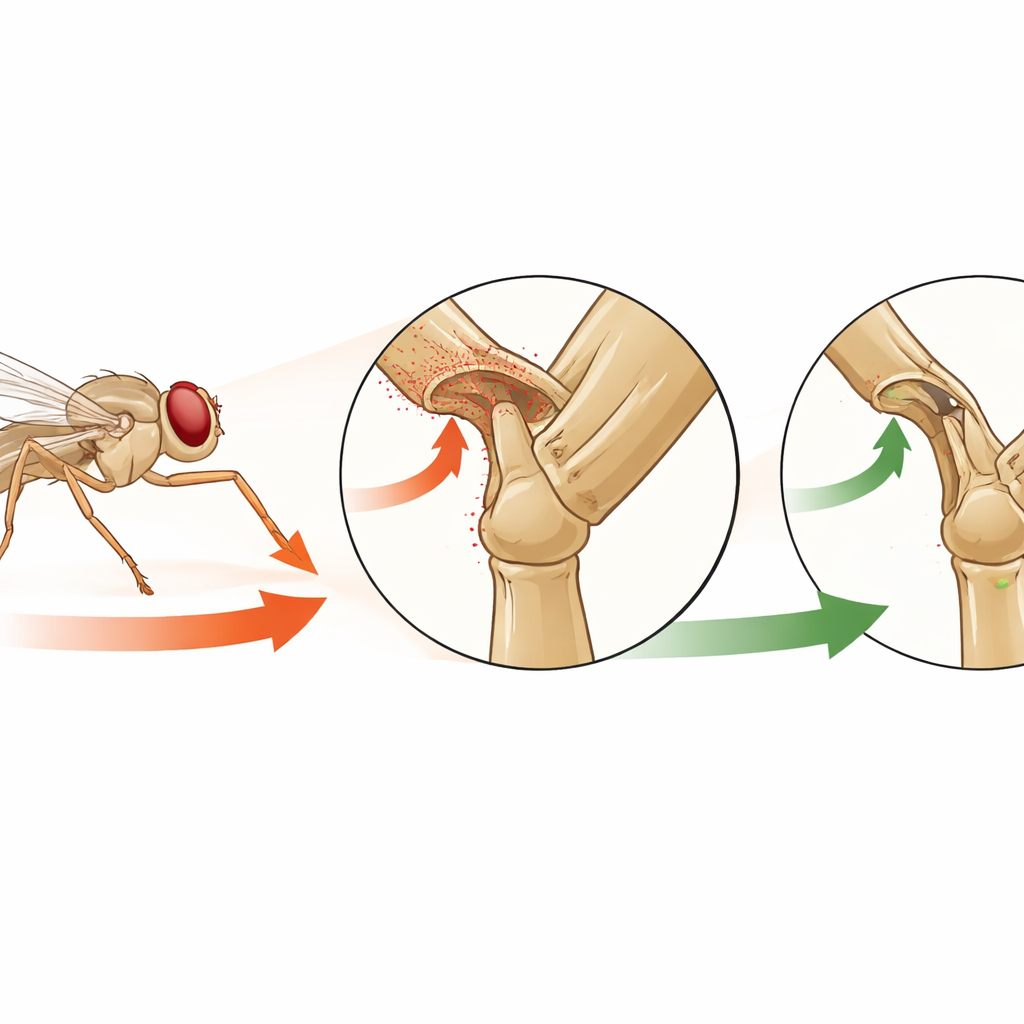
Die Autorinnen und Autoren konzentrieren sich auf eine spezielle Gruppe von berührungsähnlichen Sensoren am Bein der Fliege, sogenannte Hair Plates. Dabei handelt es sich um kleine Ansammlungen steifer Haare in Falten der Beingelenke, so angeordnet, dass sie sich nur biegen, wenn das Gelenk an den Rand seines normalen Bewegungsbereichs gedrückt wird. Mit hochauflösender Bildgebung und genetischen Werkzeugen kartierte das Team alle Hair Plates am Fliegenbein und zoomte dann auf eine bestimmte Gruppe am Vorderbein, CxHP8. Ihr Ziel war zu verstehen, welche Beinwinkel diese Sensoren detektieren, wie sie im Nervensystem verschaltet sind und welche Rolle sie bei natürlichen Verhaltensweisen spielen.
Nervenaktivität beobachten, während das Bein bewegt wird
Um zu sehen, wann CxHP8 aktiv wird, nutzten die Forschenden ein Zwei‑Photonen‑Mikroskop, um Calciumsignale in seinen Nervenfasern zu messen — eine etablierte Methode, Aktivität nachzuverfolgen. Gleichzeitig kontrollierten und verfolgten sie präzise die Position eines Vorderbeins in drei Dimensionen. Beim langsamen Bewegen des Beins zeigte sich, dass CxHP8 am stärksten feuert, wenn das obere Beinglied nach innen rotiert und zum Körper hin verschoben wird — Positionen, die dem weit nach vorn und zur Mittellinie gestreckten Vorderbein entsprechen. Wenn das Bein in diesen extremen Positionen gehalten wurde, blieb die Aktivität hoch statt nur kurz aufzuflackern, was zeigt, dass diese Sensoren kontinuierlich signalisieren, dass das Gelenk am Anschlag ist. Bei natürlichen Verhaltensweisen wie Gehen und Putzen auf einer kleinen Kugel wurden dieselben Winkel erreicht, und CxHP8 blieb aktiv, wann immer das Bein zu diesen Extremen schwang.
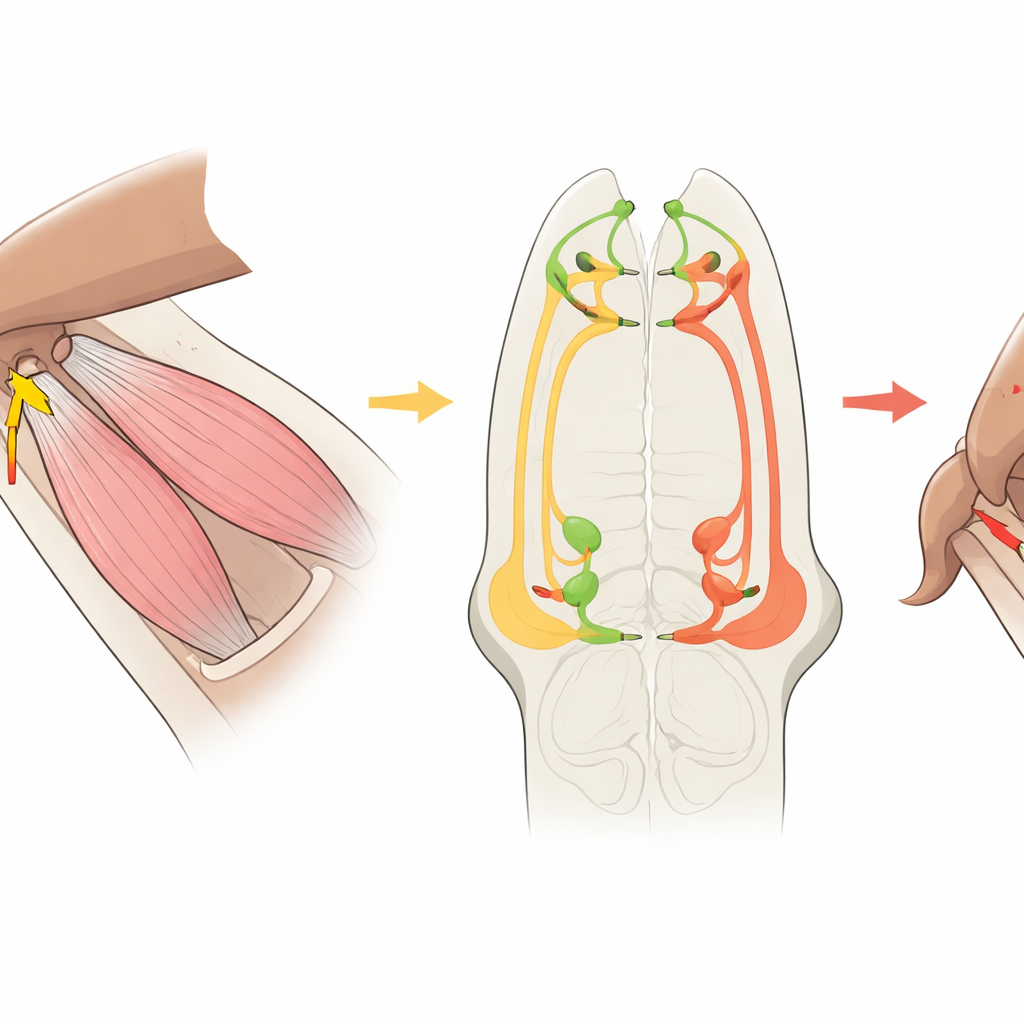
Die Verschaltung vom Sensor zum Muskel verfolgen
Das Team wandte sich dann einer detaillierten Verschaltungskarte des ventralen Nervenstrangs der Fliege zu, rekonstruiert aus Elektronenmikroskopie tausender Neuronen. Sie verfolgten die CxHP8‑Fasern vom Bein ins Nervensystem und identifizierten alle ihre Verbindungen. Die meisten Ausgänge von CxHP8 gingen direkt oder indirekt zu Beinmotorneuronen, insbesondere zu solchen, die das Bein nach hinten ziehen, sowie zu einer kleineren Gruppe, die es nach vorne schieben. In diesem Schaltkreis erregt CxHP8 stark die Motorneuronen, die das Bein rückwärts bewegen, und unterdrückt über inhibitorische Zwischenzellen Motorneuronen, die Vorwärtsbewegung antreiben. Dieses Muster legt eine einfache Regel nahe: Wenn das Bein seine Vorwärtsgrenze erreicht und CxHP8 feuert, sollte der Schaltkreis das Bein aus der Vorwärtsschwing‑Phase in eine rückwärtige, gewichtstragende Phase überführen.
Signal erzwingen und entfernen während echten Verhaltens
Um diese Vorhersage zu testen, nutzten die Autorinnen und Autoren lichtempfindliche Proteine, um CxHP8 bei verhaltenden Fliegen entweder zu aktivieren oder zu stummschalten. Wenn sie rotes Licht auf das Gelenk befestigter Fliegen mit aktivierbarem CxHP8 blitzen ließen, bewegte sich das stimulierte Bein schnell nach hinten, drehte sich nach außen und beugte sich, mit geringen Effekten auf die anderen Beine. Umgekehrt neigte das Vorderbein dazu, seine übliche Vorwärtsreichweite beim Gehen zu überschießen, wenn CxHP8 kurzzeitig mit einem lichtgesteuerten Kanal oder dauerhaft durch einen Kaliumkanal stillgelegt wurde. Sowohl auf einer schwimmenden Kugel als auch auf einem motorisierten Laufband endeten Schritte mit dem Fuß etwas weiter vorne und näher zur Mittellinie, und die Schwungphase legte eine längere Strecke zurück, obwohl das Timing der Schritte und die Koordination zwischen den Beinen weitgehend erhalten blieben. In Ruhe stellten Fliegen mit stummgeschaltetem CxHP8 ihre Beine außerdem weiter auseinander und bildeten so eine breitere Standfläche.
Viele kleine Sensoren, jeder mit einer Aufgabe
Nachdem die Funktion einer Hair Plate bestätigt war, nutzten die Forschenden dieselbe Verschaltungskarte, um die Rollen weiterer Hair Plates an benachbarten Gelenken vorherzusagen. Sie rekonstruierten deren Nervenfasern und zählten die Synapsen auf verschiedene Gruppen von Motorneuronen. Diese Analyse legt nahe, dass jede Hair Plate auf eine bestimmte Richtung des Gelenkanschlags eingestellt ist — einige zugunsten der Vorwärtsbewegung, andere der Rückwärtsbewegung und wieder andere stabilisierend dazwischen — indem sie bevorzugt die Muskeln ansteuern, die das betreffende Beinglied bewegen. Effektiv ist das Fliegenbein von kleinen, spezialisierten Grenzdetektoren umringt, die jeweils vorverdrahtet sind, das Bein von dem konkreten Extrem wegzudrücken, das sie überwachen.
Warum das für Bewegung wichtig ist
Diese Arbeit zeigt, wie ein winziges Insektenbein als Modell dienen kann, um Gelenk‑Grenzsensorik bei allen Tieren zu verstehen. Für eine Leserschaft ohne Fachhintergrund ist die zentrale Idee, dass das Nervensystem nicht nur verfolgt, wie schnell oder wie weit ein Gliedmaß bewegt wird; es hat auch dedizierte „Drahtfallen“, die feuern, wenn ein Gelenk zu nahe an den Rand kommt. Bei der Fruchtfliege signalisiert ein solcher Draht, CxHP8, kontinuierlich, wenn das Vorderbein zu weit nach vorn schwingt, und löst über einen einfachen, aber eindeutig definierten Schaltkreis einen Wechsel in einen rückwärtigen, stützenden Schritt aus und hilft, stabile Ruhehaltungen festzulegen. Da die Verschaltungsanatomie nun detailliert bekannt ist, zeigt die Studie außerdem, dass es zumindest in diesem kleinen Nervensystem möglich ist, spezifische Reflexe — etwa wann ein Bein vom Schwing‑ in den Standphasen wechselt — direkt aus dem Layout der Verbindungen zwischen Sensoren und Muskeln vorherzusagen.
Zitation: Pratt, B.G., Dallmann, C.J., Chou, G.M. et al. Proprioceptive limit detectors contribute to sensorimotor control of the Drosophila leg. Nat Commun 17, 2664 (2026). https://doi.org/10.1038/s41467-026-69333-z
Schlüsselwörter: Propriozeption, Drosophila, Fortbewegung, sensomotorische Schaltkreise, Hair‑Plate‑Neuronen