Clear Sky Science · ar
البنية والوظيفة لمجمع SEAC–EGOC الفائق الاستشعار للأحماض الأمينية في الخميرة
كيف تقرر الخلايا متى تنمو
يجب على كل خلية أن تقرر باستمرار ما إذا كان بإمكانها تحمل النمو أم يجب عليها الحفاظ على الموارد. يعتمد هذا القرار بشكل كبير على الأحماض الأمينية، مكوّنات البروتينات ومصدر رئيسي للنيتروجين. تكشف هذه الورقة، بتفاصيل هيكلية رائعة، كيف تستشعر خلايا الخميرة توفر الأحماض الأمينية عبر تجمّع جزيئي ضخم وتستخدم تلك المعلومات لتشغيل أو إيقاف متحكم نمو رئيسي يسمى TORC1. ونظراً لأن الدارات المماثلة محفوظة في الخلايا البشرية ومرتبطة بالسرطان والتمثيل الغذائي، فالفهم المستمد من الخميرة يقدِّم نافذة على كيفية تقييم خلايانا متى تكون الظروف مناسبة للنمو.
محرّك معقّد على مركز إعادة التدوير الخلوي
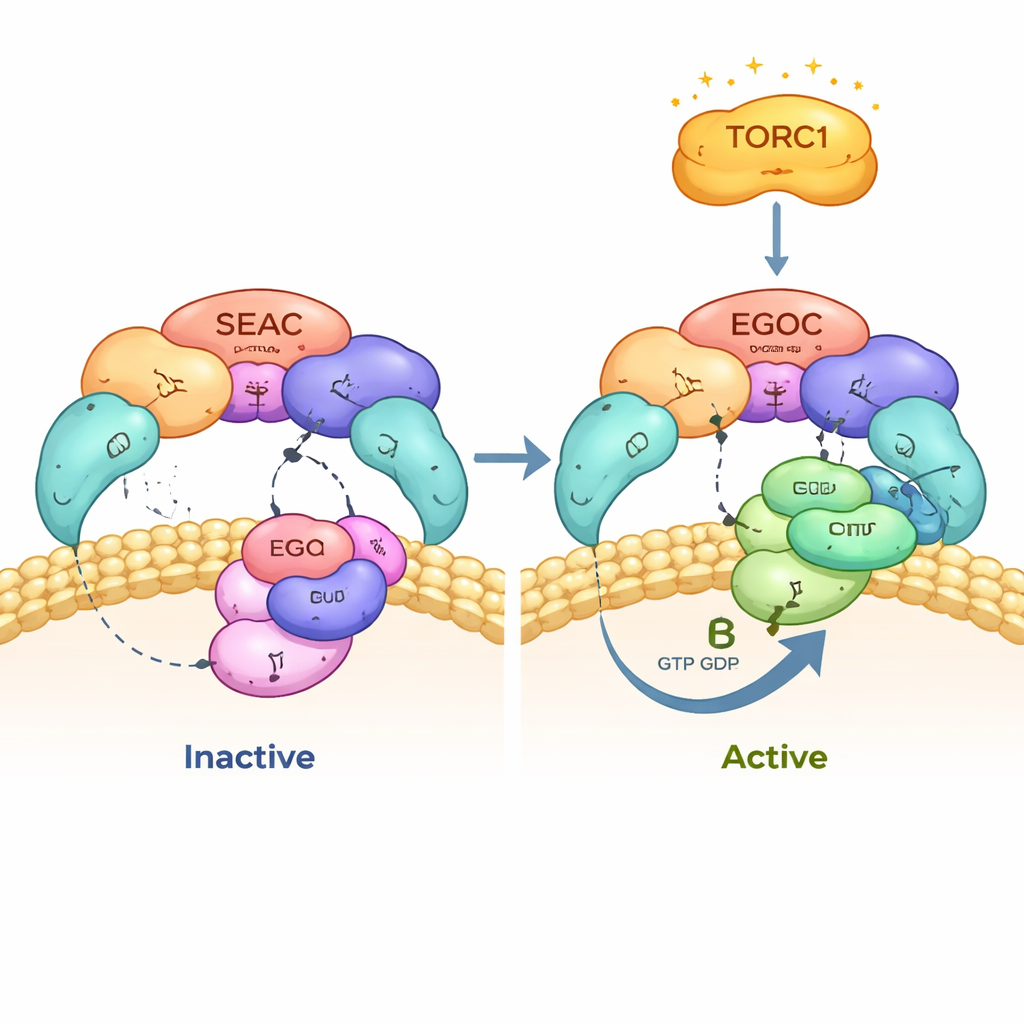
في الخميرة، يجلس مجمع مركزي لاتخاذ القرار يُدعى SEAC على غشاء الفجوة، العضية التي تعمل كمركز لإعادة التدوير داخل الخلية. هناك يلتقي بمجمع آخر، EGOC، الذي ينقل معلومات حول مستويات الأحماض الأمينية. معاً يمرران الإشارات إلى TORC1، مفتاح التحكم في النمو. ينقسم SEAC وظيفياً إلى نصفين: SEACIT القادر على إيقاف TORC1، وSEACAT الذي كان يُعتقد لفترة طويلة أنه يكبح SEACIT. استخدم المؤلفون المجهر الإلكتروني بالتبريد عالي الدقة لتصوير SEAC الكامل متصلاً بـEGOC، موثقين لقطة لهذا المحور الجزيئي تماماً في موضع حدوث الإشارة، على سطح الفجوة. 
رؤية البنية الجزيئية عن قرب
تكشف البنية أن SEAC عبارة عن لب كبير مكوّن من وحدات SEACAT، تنبثق منه "جناحان" مرنان من SEACIT. يمكن لكل جناح أن يرتبط بمجمع EGOC واحد، ما يعني أن SEAC المفرد يمكنه استقبال مدخلين إشاريين في آن واحد. وبشكل لافت، يرتبط EGOC بالأجنحة فقط وليس باللب، وشكل SEAC العام يتغير بالكاد عند ارتباط EGOC. هذا يدل على أن SEACAT لا يغلق مكان الفعالية الخاص بـSEACIT بشكل مادي ولا يمنع ارتباط EGOC. كما أن اتجاهية المجمع على غشاء الفجوة تضع لب SEAC مواجهة للسيتوبلازم، حيث يمكنها التفاعل مع منظمات إضافية، بينما تكاد الأجنحة تكون قريبة من الغشاء، في موقع مثالي للتماس مع EGOC وشركاء مثبتين على الغشاء.
كيف يختار SEAC الإشارة الصحيحة
اقترب المؤلفون من نقاط التماس بين SEAC وEGOC ووجدوا أن SEAC يتعرف فقط على النسخة "النشطة" من EGOC، والتي تكون فيها إحدى بروتينات GTPase الصغيرة فيه، Gtr1، حاملةً نوكليوتيد شبيه بـGTP، بينما شريكها Gtr2 يحمل GDP. يوفر SEACIT "إصبعاً" تحفيزياً يصل إلى جيب النوكليوتيد في Gtr1 ويسرّع تحلل GTP، محولاً EGOC من حالة نشطة إلى حالة غير نشطة. تبين خرائط الكريو-EM التفصيلية وتجارب الطفرات أن هذا الإصبع التحفيزي وبقايا مساعدة مجاورة ضروريان للتفاعل وللقراءة الصحيحة لحالة النوكليوتيد. عندما غيّر المؤلفون هذه البقايا الأساسية، لم يعد SEAC يتموضع بشكل صحيح على الفجوة، ولم يستطع EGOC الانخراط بشكل منتج.
التحكم السريع مقابل البطيء لمفتاح النمو
لفهم ما يعنيه ذلك للخلية الحية، تابع الباحثون نشاط TORC1 مع مرور الوقت عند إزالة الأحماض الأمينية ثم إعادتها. في الخلايا الطبيعية، ينطفئ TORC1 بسرعة خلال بضع دقائق من مجاعة النيتروجين ثم يعاد تشغيله بسرعة عند عودة الأحماض الأمينية. عندما تعطّل الإصبع التحفيزي في SEACIT أو حُذفت وحدات SEACIT، لم يستجب TORC1 تقريباً خلال هذه الفترة الزمنية المبكرة. وكان الأمر نفسه عندما أزيل كل من Gtr1 وGtr2، مما يبيّن أن زوج SEAC–EGOC هو المسار السريع المهيمن لتحكم الأحماض الأمينية في TORC1. إلا أنه لاحقاً—بعد نحو نصف ساعة—تكيَّف نشاط TORC1 ببطء حتى في هذه الطفرات، مما يشير إلى أن مساراً منفصلاً وبطيئاً نحو TORC1، مستقل عن نظام GTPase هذا، يعمل أيضاً في الخلايا.
دور مفاجئ لوحدة بعيدة
من النتائج المثيرة للاهتمام ما يتعلق بـSea2، مكوّن من SEACAT يملك نطاقاً مرناً شبيهاً بالمروحة يبرز من اللب. أدى إزالتها، أو مجرد إزالة مروحتها، إلى عيوب إشارات تقارب إلى حد كبير تلك التي تُرى عند فقدان النشاط التحفيزي في SEAC: استجاب TORC1 ببطء لكل من سحب الأحماض الأمينية وإعادتها. ومع ذلك، يقع Sea2 بعيداً عن الموقع التحفيزي ولا يلامس جناح SEACIT مباشرة، ما يجعل تفسيراً بسيطاً يقوم على الحجب غير مرجح. أشارت تجارب جينية جمعت فقدان Sea2 مع نسخة SEAC خاملة تحفيزياً إلى أن قفل نشاط GAP كاملاً أو إيقافه تماماً يعيق الاستجابة السريعة، بينما يلزم تنظيم متوسط. يقترح المؤلفون أن مروحة Sea2 تعمل كمنصة ارتكاز لعامل مجهول يهدئ نشاط SEACIT، مما يسمح للخلايا بضبط TORC1 بدقة بدلاً من إيقافه نهائياً.
ما يعنيه هذا لقرارات نمو الخلايا
بشكل عام، تعيد الدراسة صياغة SEAC ليس كمجمّعين متعارضين بل كآلة متكاملة تعمل فيها الجناح التحفيزي (SEACIT) واللب البنيوي (SEACAT) معاً لاستشعار الأحماض الأمينية وتعديل مفتاح النمو TORC1 بسرعة. يوفر الفعل الأنزيمي لـSEACIT على EGOC التحكم السريع والدقيق، بينما ربما تستقطب مروحة Sea2 منظمات تعدل هذا النشاط. في الوقت نفسه، تضمن مسارات موازية أبطأ أن TORC1 يعكس حالة المغذيات في نهاية المطاف حتى إذا تعطّل جهاز الاستشعار الأساسي. وبما أن مجمعات قريبة الصلة تقوم بنفس الوظيفة في الخلايا البشرية، فإن هذه النتائج من الخميرة توضح كيفية موازنة خلايانا للنمو مع توافر المغذيات، مع تداعيات لفهم الأمراض التي يختل فيها إشعار TORC1.
الاستشهاد: Tafur, L., Bonadei, L., Zheng, Y. et al. Structure and function of the yeast amino acid-sensing SEAC–EGOC supercomplex. Nat Struct Mol Biol 33, 488–498 (2026). https://doi.org/10.1038/s41594-026-01746-2
الكلمات المفتاحية: استشعار الأحماض الأمينية, إشارات TORC1, مجمع SEAC GATOR, ضبط نمو الخلايا, المجهر الإلكتروني بالتبريد